Gemensam härkomst
| Del av en serie om |
| Evolutionsbiologi |
|---|

|
Gemensam härkomst är ett begrepp inom evolutionsbiologin som är tillämpligt när en art är förfader till två eller flera arter senare i tiden. Enligt modern evolutionär biologi kan alla levande varelser vara ättlingar till en unik förfader som vanligtvis kallas den sista universella gemensamma förfadern (LUCA) av allt liv på jorden .
Vanlig härkomst är en effekt av artbildning , där flera arter härstammar från en enda förfäderspopulation. Ju nyare urbefolkningen två arter har gemensamt, desto närmare är de släkt. Den senaste gemensamma förfadern av alla för närvarande levande organismer är den sista universella förfadern, som levde för cirka 3,9 miljarder år sedan . De två tidigaste bevisen för liv på jorden är grafit som befunnits vara biogen i 3,7 miljarder år gamla metasedimentära bergarter som upptäckts i västra Grönland och mikrobiella mattfossiler som hittats i 3,48 miljarder år gammal sandsten upptäckt i västra Australien . Alla för närvarande levande organismer på jorden delar ett gemensamt genetiskt arv, även om förslaget om betydande horisontell genöverföring under tidig evolution har lett till frågor om livets monophyly (enda härkomst). 6 331 grupper av gener som är gemensamma för alla levande djur har identifierats; dessa kan ha uppstått från en enda gemensam förfader som levde för 650 miljoner år sedan i prekambrium .
Universell gemensam härkomst genom en evolutionär process föreslogs först av den brittiske naturforskaren Charles Darwin i den avslutande meningen i hans bok från 1859 On the Origin of Species :
Det finns storhet i denna livsåskådning, med dess flera krafter, som ursprungligen har andats i några få former eller i en; och att medan denna planet har cyklat vidare enligt tyngdlagens fasta lag, från en så enkel början ändlösa former har de vackraste och mest underbara utvecklats och håller på att utvecklas.
Historia
Tanken att alla levande varelser (inklusive saker som anses icke-levande av vetenskapen) är relaterade är ett återkommande tema i många inhemska världsbilder över hela världen. Senare, på 1740-talet, kom den franske matematikern Pierre Louis Maupertuis fram till idén att alla organismer hade en gemensam förfader och hade divergerat genom slumpmässig variation och naturligt urval . I Essai de cosmologie (1750) noterade Maupertuis:
Får vi inte säga att i den slumpmässiga kombinationen av naturens produktioner, eftersom endast de varelser kunde överleva i vars organisationer en viss grad av anpassning fanns, finns det inget extraordinärt i det faktum att sådan anpassning faktiskt finns i alla dessa arter som finns nu? Slumpen, kan man säga, visade sig vara ett stort antal individer; en liten del av dessa var organiserade på ett sådant sätt att djurens organ kunde tillgodose deras behov. Ett mycket större antal visade varken anpassning eller ordning; dessa sistnämnda har alla gått under... De arter som vi ser idag är alltså bara en liten del av alla de som ett blindt öde har frambringat.
skrev filosofen Immanuel Kant i Kritik der Urteilskraft ( Kritik av omdömet ) att djurformernas likhet innebär en gemensam ursprungstyp, och därmed en gemensam förälder.
1794 frågade Charles Darwins farfar, Erasmus Darwin :
Skulle det vara för djärvt att föreställa sig att det under den långa tiden, sedan jorden började existera, kanske miljoner åldrar innan mänsklighetens historia började, skulle det vara för djärvt att föreställa sig att allt var varmt -blodiga djur har uppstått ur en levande glödtråd, som den stora Första Orsaken gavs med djurlighet, med kraften att förvärva nya delar åtföljda av nya benägenheter, styrda av irritationer, förnimmelser, viljor och associationer; och därmed besitta förmågan att fortsätta att förbättras genom sin egen inneboende verksamhet, och att leverera ned dessa förbättringar i generation till sina efterkommande, en värld utan slut?
Charles Darwins åsikter om gemensam härkomst, som de uttrycktes i On the Origin of Species , var att det var troligt att det bara fanns en stamfader för alla livsformer:
Därför bör jag dra slutsatsen från analogi att förmodligen alla de organiska varelser som någonsin har levt på denna jord har härstammat från någon urform, i vilken liv först andades in.
Men han föregår det påpekandet med: "Analogi skulle leda mig ett steg längre, nämligen till tron att alla djur och växter har härstammat från någon prototyp. Men analogi kan vara en bedräglig vägledning." Och i den efterföljande upplagan hävdar han snarare,
"Vi känner inte till alla möjliga övergångsgraderingar mellan de enklaste och de mest perfekta organen; det kan inte låtsas att vi känner till alla de olika distributionssätten under de långa åren, eller att vi vet hur ofullkomlig den geologiska journalen är. Hur allvarliga dessa svårigheter än är, enligt min bedömning störtar de inte teorin om härkomst från ett fåtal skapade former med efterföljande modifiering."
Gemensam härkomst var allmänt accepterad bland det vetenskapliga samfundet efter Darwins publicering. År 1907 Vernon Kellogg att "praktiskt taget inga naturforskare av position och erkänd prestation tvivlar på teorin om härkomst."
2008 noterade biologen T. Ryan Gregory att:
Ingen tillförlitlig observation har någonsin hittats som motsäger den allmänna uppfattningen om gemensam härkomst. Det borde därför inte komma som någon överraskning att det vetenskapliga samfundet i stort har accepterat evolutionär härkomst som en historisk verklighet sedan Darwins tid och anser att det är bland de mest tillförlitligt etablerade och fundamentalt viktiga fakta inom all vetenskap.
Bevis
Vanlig biokemi
Alla kända livsformer är baserade på samma grundläggande biokemiska organisation: genetisk information kodad i DNA , transkriberad till RNA , genom effekten av protein- och RNA- enzymer , sedan översatt till proteiner av (mycket liknande) ribosomer , med ATP , NADPH och andra som energikällor. Analys av små sekvensskillnader i vitt delade substanser såsom cytokrom c stöder ytterligare universell gemensam härkomst. Cirka 23 proteiner finns i alla organismer och fungerar som enzymer som utför kärnfunktioner som DNA-replikation. Det faktum att endast en sådan uppsättning enzymer existerar är övertygande bevis på en enda härkomst. 6 331 gener som är gemensamma för alla levande djur har identifierats; dessa kan ha uppstått från en enda gemensam förfader som levde för 650 miljoner år sedan i prekambrium .
Vanlig genetisk kod
| Aminosyror | opolär | polär | grundläggande | surt | Stoppa kodon |
|
1: a basen |
2:a basen | |||||||
|---|---|---|---|---|---|---|---|---|
| T | C | A | G | |||||
| T | TTT |
Fenylalanin _ |
TCT | Serine | TAT | Tyrosin | TGT | Cystein |
| TTC | TCC | TAC | TGC | |||||
| TTA | Leucin | TCA | TAA | Sluta | TGA | Sluta | ||
| TTG | TCG | MÄRKA | Sluta | TGG | Tryptofan | |||
| C | CTT | CCT | Proline | KATT | Histidin | CGT | Arginin | |
| CTC | CCC | CAC | CGC | |||||
| CTA | CCA | CAA | Glutamin | CGA | ||||
| CTG | CCG | CAG | CGG | |||||
| A | ATT | Isoleucin | SPELA TEATER | Treonin | AAT | Asparagin | AGT | Serine |
| ATC | ACC | AAC | AGC | |||||
| ATA | ACA | AAA | Lysin | AGA | Arginin | |||
| ATG | Metionin | ACG | AAG | AGG | ||||
| G | GTT | Valine | GCT | Alanin | GAT |
Asparaginsyra _ |
GGT | Glycin |
| GTC | GCC | GAC | GGC | |||||
| GTA | GCA | GAA |
Glutaminsyra _ |
GGA | ||||
| GTG | GCG | GAG | GGG | |||||
Den genetiska koden ("översättningstabellen" enligt vilken DNA-information översätts till aminosyror , och därmed proteiner) är nästan identisk för alla kända livsformer, från bakterier och arkéer till djur och växter . Allmänheten i denna kod anses allmänt av biologer som ett definitivt bevis för universell gemensam härkomst.
Sättet som kodon (DNA-tripletter) kartläggs till aminosyror verkar vara starkt optimerat. Richard Egel hävdar att särskilt de hydrofoba (icke-polära) sidokedjorna är välorganiserade, vilket tyder på att dessa gjorde det möjligt för de tidigaste organismerna att skapa peptider med vattenavstötande regioner som kan stödja de väsentliga elektronbytesreaktionerna ( redox ) för energiöverföring.
Selektivt neutrala likheter
Likheter som inte har någon adaptiv relevans kan inte förklaras av konvergent evolution , och därför ger de övertygande stöd för universell gemensam härkomst. Sådana bevis har kommit från två områden: aminosyrasekvenser och DNA-sekvenser. Proteiner med samma tredimensionella struktur behöver inte ha identiska aminosyrasekvenser; alla irrelevanta likheter mellan sekvenserna är bevis för gemensam härkomst. I vissa fall finns det flera kodon (DNA-tripletter) som kodar redundant för samma aminosyra. Eftersom många arter använder samma kodon på samma plats för att specificera en aminosyra som kan representeras av mer än ett kodon, är det bevis för att de delar en nyligen gemensam förfader. Hade aminosyrasekvenserna kommit från olika förfäder, skulle de ha kodats för av vilket som helst av de överflödiga kodonen, och eftersom de korrekta aminosyrorna redan skulle ha funnits, skulle det naturliga urvalet inte ha drivit fram någon förändring i kodonen, hur mycket som helst. tid fanns tillgänglig. Genetisk drift kan förändra kodonen, men det skulle vara extremt osannolikt att alla överflödiga kodoner i en hel sekvens matchar exakt över flera linjer. På liknande sätt ger delade nukleotidsekvenser, särskilt där dessa uppenbarligen är neutrala, såsom placeringen av introner och pseudogener , starka bevis på gemensam härkomst.
Andra likheter
Biologer [ kvantifierar ] pekar ofta på universaliteten hos många aspekter av cellulärt liv som stödjande bevis för de mer övertygande bevis som anges ovan. Dessa likheter inkluderar energibäraren adenosintrifosfat (ATP), och det faktum att alla aminosyror som finns i proteiner är vänsterhänta . Det är dock möjligt att dessa likheter resulterade på grund av fysikens och kemins lagar - snarare än genom universell gemensam härkomst - och därför resulterade i konvergent evolution. Däremot finns det bevis för homologi av de centrala subenheterna av transmembrana ATPaser i alla levande organismer, särskilt hur de roterande elementen är bundna till membranet. Detta stöder antagandet om en LUCA som en cellulär organism, även om primordiala membran kan ha varit semipermeabla och utvecklats senare till membranen hos moderna bakterier, och på en andra väg till de hos moderna archaea också.
Filogenetiska träd
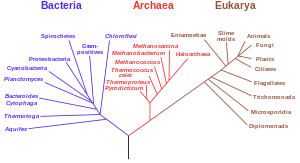
Ett annat viktigt bevis är från detaljerade fylogenetiska träd (dvs. "genealogiska träd" av arter) som kartlägger de föreslagna indelningarna och gemensamma förfäder för alla levande arter. 2010 publicerade Douglas L. Theobald en statistisk analys av tillgängliga genetiska data, som kartlade dem till fylogenetiska träd, som gav "starkt kvantitativt stöd, genom ett formellt test, för livets enhet."
Traditionellt har dessa träd byggts med hjälp av morfologiska metoder, såsom utseende, embryologi , etc. På senare tid har det varit möjligt att konstruera dessa träd med hjälp av molekylär data, baserat på likheter och skillnader mellan genetiska sekvenser och proteinsekvenser. Alla dessa metoder ger i huvudsak liknande resultat, även om de flesta genetiska variationer inte har något inflytande över extern morfologi. Att fylogenetiska träd baserade på olika typer av information överensstämmer med varandra är ett starkt bevis på en verklig underliggande gemensam härkomst.
Invändningar
Genutbytesmoln fylogenetisk analys
Theobald noterade att betydande horisontell genöverföring kunde ha inträffat under tidig evolution. Bakterier förblir idag kapabla till genutbyte mellan avlägset besläktade linjer. Detta försvagar det grundläggande antagandet om fylogenetisk analys, att likheten mellan genomer innebär gemensamma anor, eftersom tillräckligt genutbyte skulle tillåta linjer att dela mycket av sitt genom oavsett om de delade en förfader (monofyl) . Detta har lett till frågor om livets enda härkomst. Biologer anser dock att det är mycket osannolikt att helt obesläktade protoorganismer kan ha utbytt gener, eftersom deras olika kodningsmekanismer endast skulle ha resulterat i förvanskning snarare än fungerande system. Senare kunde dock många organismer som alla härrörde från en enda förfader lätt ha delade gener som alla fungerade på samma sätt, och det verkar som att de har det.
Konvergent evolution
Om tidiga organismer hade drivits av samma miljöförhållanden för att utveckla liknande biokemi konvergent, skulle de självständigt kunna ha förvärvat liknande genetiska sekvenser. Theobalds "formella test" kritiserades följaktligen av Takahiro Yonezawa och kollegor för att inte ta hänsyn till konvergens. De hävdade att Theobalds test var otillräckligt för att skilja mellan de konkurrerande hypoteserna. Theobald har försvarat sin metod mot detta påstående och hävdat att hans tester skiljer mellan fylogenetisk struktur och ren sekvenslikhet. Därför, hävdade Theobald, visar hans resultat att "riktiga universellt konserverade proteiner är homologa ."
RNA-världen
Möjligheten nämns ovan att alla levande organismer kan härstamma från en ursprunglig encellig organism med ett DNA- genom , och att detta innebär ett enda ursprung för livet. Även om en sådan universell gemensam förfader kan ha funnits, är det osannolikt att en sådan komplex enhet har uppstått spontant från icke-liv och därför kan en cell med ett DNA-genom rimligen inte betraktas som livets "ursprung". För att förstå livets "ursprung" har det föreslagits att DNA-baserat cellulärt liv härstammar från relativt enkla precellulära självreplikerande RNA - molekyler som kan genomgå naturligt urval (se RNA-världen ). Under evolutionens gång ersattes denna RNA-värld av den evolutionära framväxten av DNA-världen. En värld av självständigt självreplikerande RNA-genom existerar tydligen inte längre (RNA-virus är beroende av värdceller med DNA-genom). Eftersom RNA-världen uppenbarligen är borta, är det inte klart hur vetenskapliga bevis skulle kunna användas på frågan om det fanns en enda "ursprung" till livshändelse som allt liv härstammar från.
Se även
Bibliografi
- Crombie, AC ; Hoskin, Michael [på slovakiska] (1970). "Den vetenskapliga rörelsen och spridningen av vetenskapliga idéer, 1688–1751". I Bromley, JS (red.). Storbritanniens och Rysslands uppkomst, 1688–1715/25 . Den nya Cambridge moderna historien . Vol. 6. London: Cambridge University Press . ISBN 978-0-521-07524-4 . LCCN 57014935 . OCLC 7588392 .
- Darwin, Charles (1859). Om arternas ursprung med hjälp av naturligt urval, eller bevarandet av gynnade raser i kampen för livet ( 1:a uppl.). London: John Murray . LCCN 06017473 . OCLC 741260650 . Boken är tillgänglig från The Complete Work of Charles Darwin Online . Hämtad 2015-11-23.
- Darwin, Erasmus (1818) [Ursprungligen publicerad 1794]. Zoonomi; eller lagarna för organiskt liv . Vol. 1 (4:e amerikanska upplagan). Philadelphia, PA: Edward Earle. Zoonomi; eller The laws of organic life: in three parts (Volume 1) (1818) på Internet Archive Retrieved 2015-11-23.
- Harris, C. Leon (1981). Evolution: Genesis and Revelations: With Readings from Empedocles to Wilson . Albany, NY: State University of New York Press . ISBN 978-0-87395-487-7 . LCCN 81002555 . OCLC 7278190 .
- Kant, Immanuel (1987) [Ursprungligen publicerad 1790 i Preussen som Kritik der Urteilskraft ]. Kritik av domen . Översatt, med en inledning, av Werner S. Pluhar; förord av Mary J. Gregor . Indianapolis, IN: Hackett Publishing Company . ISBN 978-0-87220-025-8 . LCCN 86014852 . OCLC 13796153 .
- Treasure, Geoffrey (1985). Skapandet av det moderna Europa, 1648-1780 . New York: Methuen . ISBN 978-0-416-72370-0 . LCCN 85000255 . OCLC 11623262 .
- Ziegler, Heinrich Ernst (1904). Der Begriff des Instinktes einst und jetzt . Jena.
Anteckningar
externa länkar
- 29+ Evidences for Macroevolution: The Scientific Case for Common Descent från TalkOrigins Archive .
- Livets träd webbprojekt

{kind=link}
.svg){kind=link}