JADE1
| JADE1 | |||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Identifiers | |||||||||||||||||||||||||||||||||||||||||||||||
| , PHF17, jade familj PHD finger 1 | |||||||||||||||||||||||||||||||||||||||||||||||
| Externa ID:n | |||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||
| Wikidata | |||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||







JADE1 är ett protein som hos människor kodas av JADE1 -genen .
Familj
En liten familj av proteiner som heter Gene for Apoptosis and Differentiation (JADE) inkluderar tre medlemmar som kodas av individuella gener: Plant Homeo-domän-17 (PHF17, JADE1), PHF16 (JADE3) och PHF15 (JADE2). Alla proteiner i JADE-familjen bär två anmärkningsvärda mellanmolekyldomäner: det kanoniska Plant Homeo-domänen (PHD) zinkfinger och utökat PHD-liknande zinkfinger. JADE1 klassificeras därför som en medlem av PHD-proteinfamiljen. Det finns två kända proteinprodukter av PHF17-genen, JADE1 i full längd (JADE1L) och dess splitsningsvariant som saknar det C-terminala fragmentet som också kallas kort isoform (JADE1S).
Upptäckt
Nagase et al. klonade och sekvenserade 100 individuella cDNA från fetala hjärnans cDNA-bibliotek, inklusive klonen KIAA1807 som betecknades PHF17. Den förutsagda proteinprodukten på 702 aminosyror från den klonen liknade det humana zinkfingerproteinet BR140 (BRPF1). Baserat på sekvensdatabasanalys föreslog studien att PHF17 kan fungera i nukleinsyrahanteringsvägar. Genom att använda jäst två hybrid pull-down-metod för att söka efter nya partner till proteinprodukten från Von Hippel Lindau-genen (pVHL) identifierade en annan studie cDNA som matchade KIAA1807-klonen. Proteinprodukten av detta cDNA fick namnet JADE1 (Jade-1, PHF17). Den härledda 509 aminosyror långa proteinprodukten av JADE1-cDNA bekräftades ytterligare som fysisk partner till pVHL. I en genetisk screeningstudie som sökte efter gener involverade i embryogenes, identifierades musortologen av JADE1. Den studien gav den första karakteriseringen av JADE1-genen och definierade en ny JADE-familj. Studien gav möss med knock-out av JADE1-genen.
Jade1-transkript hos både människor och möss genomgår alternativ splitsning och polyadenylering som producerar två huvudtranskript, fullängds 6 kb mRNA och 3,6 kb mRNA. Två resulterande proteinprodukter av JADE1-genen betecknades JADE1S för den korta (vilket är samma som (3)) och JADE1L för den långa isoformen. Flera mindre avskrifter påvisas också. Databasanalysen avslöjade ytterligare två JADE1-paraloger och medlemmar av JADE-familjen, JADE2 och JADE3. JADE3 är identisk med E9-protein som identifierats i en tidigare oberoende studie som föreslog roll i apoptos för PHF16/JADE3/E9 i bröstcancerceller.
JADE1 har kartlagts till kromosom 4 (4q26-q27). JADE1 är bevarad och dess ortologer har hittats eller förutspåtts i nästan varje metazoan. Genstruktur och sekvenser, varianter, konservering, ortologer och paraloger, JADE1 fylogenetiska träd och storskalig screening av JADE1-vävnadsuttryck finns i flera omfattande databaser ( https://www.genecards.org ; http://useast.ensembl. org ).
Strukturera
JADE1-polypeptiden i full längd har en kanonisk och en utökad PHD-zinkfingerdomän. Andra domäner inkluderar den N-terminala kandidat-PEST-domänen, Enhancer of polycomb-liknande domän och den C-terminala nukleära lokaliseringssignalen (NLS) (prosite.expasy.org). JADE1-protein är ett mål för posttranslationella modifieringar, inklusive fosforylering (Fig 1)
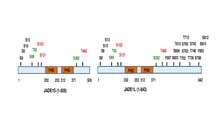
). Sex aminosyrarester identifierades vara fosforylerade på cellcykelberoende sätt via Aurora A-kinasväg . JADE1 är ett mål för fosforylering av kaseinkinas 2 (CK2). Dessutom hittas flera fosforyleringsställen genom screeningmetoder med hög genomströmning och i silicoanalys. Sammanfattningsschema för JADE1L och JADE1S proteiner fosforyleringsställen med referenser finns i.
Proteiner som bär i tandem kanoniska och utökade PHD-fingrar bildar en liten underfamilj inom den stora PHD-proteinfamiljen (www.genenames.org). Andra proteiner som bär tandem av PHD-fingrar och relaterade till JADE1 inkluderar proteiner som är komponenter i kromatinbindande och modifierande komplex BRPF1, BRPF3, BRD1. Kristallstrukturen för JADE1 PHD-domäner har inte lösts. Kanoniskt PHD-fingermotiv har signaturen C4HC3, representerar relativt liten, stabil struktur och skiljer sig från RINGfingret av typen C3HC4. PHD-domäner kan känna igen och binda specifikt metylerat lysin av histon H3, vilket definierade dessa domäner som epigenetiska histonkodläsare. Det finns recensioner som beskriver struktur och egenskaper hos PHD-fingrar på djupet.
Cellulär funktion
JADE1-proteiner är multifunktionella och interagerar med flera proteinpartners.
Histonacetylering
Funktion av JADE1 i histonacetylering och transkriptionsaktivering som krävde det andra förlängda PHD-zinkfingret rapporterades 2004(16). JADE1 ökar dramatiskt nivåerna av acetylerad histon H4 i kromatin, men inte histon H3, en specificitet som är karakteristisk för MYST-familjen HAT TIP60 och HBO1. TIP60 associerar fysiskt med JADE1 och förstärker JADE1 HAT-funktionen i levande celler. TIP60 och JADE1 stabiliserade varandra ömsesidigt. Transkriptions- och HAT-aktiviteter för JADE1 kräver PHD2. Resultaten tyder på kromatininriktningsrollen för JADE1 PHD2. Dessutom binder PHD2 av JADE1 den N-terminala svansen av histon H3 inom kromatinsammanhang oavsett metyleringsstatus.
Studier som analyserar inhemska komplex av Hämmare av tillväxt (ING) PHD fingerfamilj av proteiner avslöjade att ING4- och ING5-proteiner är associerade med JADE1S och HAT HBO1, medan ING3 är associerad med EPC1 (JADE1-homolog), TIP60 (HBO1-homolog) och flera andra partners . Båda komplexen inkluderade också ett litet Eaf6-protein. Den biokemiska analysen och i kiselanalys av komplex bildade av HBO1 och TIP60 föreslog gemensam arkitektur och stödde rollen för JADE1 i bulkhiston H4-acetylering. Karakterisering av JADE1 och HBO1 funktionella interaktioner visar strukturella och funktionella likheter mellan komplexen(16, 19). På samma sätt som TIP60 stabiliserar JADE1 och HBO1 varandra. JADE1 binder till och gör det möjligt för HBO1 att förbättra global histon H4-acetylering, vilket kräver intakt PHD2-finger. På samma sätt som HBO1 är JADE1 ansvarig för bulk histon H4 acetylering i odlade celler. H4K5, H4K12 och troligen H4K8 är mål för JADE1-beroende acetylering i odlade celler och in vivo. Flera potentiella transkriptionsmål för JADE1 har föreslagits från experiment med screeningmetoder. Enligt screening genomisk analys gjord av ChIP-chip-analysen återfinns JADE1L-komplexet huvudsakligen längs de kodande regionerna för många gener och överflöd av JADE1L korrelerar mestadels med H3K36me3 -histonmärket. JADE1L överuttryck korrelerar med ökade mängder H4acK8 i den kodande regionen av många gener. De två PHD-zinkfingrarna hos JADE1 verkar binda företrädesvis icke-metylerad N-terminal peptid av histon. JADE1 isoformer sätter samman minst två olika komplex, JADE1L-HBO1-ING4/5 och JADE1S-HBO1 komplex. På grund av avsaknaden av det C-terminala fragmentet är JADE1S oförmögen att binda ING4/5-partners. Ett litet mindre karakteriserat protein Eaf6 är också en annan komponent i JADE1-komplex.
Cellcykel
Acetylering av N-terminala fragment av bulkhiston H4 har varit känt för att korrelera med DNA-syntes och celldelning. Flera studier stödjer cellcykelrollen för JADE1 kopplat till HBO1-vägen. Både JADE1 och HBO1 krävs oberoende för acetylering av bulkhiston H4 i odlade celler. Utarmningen av JADE1-proteiner av siRNA resulterar i 1) minskade nivåer av histon H4 bulk-acetylering; 2) långsammare hastigheter för DNA-syntes i odlade celler; 3) minskade nivåer av total och kromatinbunden HBO1; 4) upphävande av kromatinrekrytering av MCM7. Håller med om dessa resultat, JADE1L-överuttryck ökar kromatinbundet MCM3-protein. Effekterna av JADE1-utarmning på DNA-replikationshändelser liknar de som ursprungligen beskrevs för HBO1 och antyder adapterroll för JADE1 i HBO1-medierad cellcykelreglering.
JADE1 roll i DNA-skada har föreslagits. Ett nyligen upptäckt icke-kodande RNA lncRNA-JADE reglerar JADE1-uttryck och ger en funktionell koppling mellan DNA-skaderespons (DDR) och bulkhiston H4-acetylering. Resultat stödjer roll i DNA-syntes kopplad till histon H4-acetylering. I odlade celler slår ner lncRNA-JADE ökad cellkänslighet för DNA-skadande läkemedel. I möss tumör xenograft modell, knock down av lncRNA-JADE hämmade xenograft brösttumör tillväxt. I en human pilotstudie upptäcktes högre nivåer av lncRNA-JADE såväl som JADE1-protein i bröstcancervävnader jämfört med normala vävnader. Slutligen korrelerade de högre nivåerna av JADE1-protein omvänt med överlevnadsfrekvensen för patienter med bröstcancer. Studien tyder på att lncRNA-JADE kan bidra till brösttumörbildning, och att JADE1-protein förmedlar åtminstone en del av denna effekt. JADE1 och cytokines. JADE1S reglerar negativt cytokines av epitelcellscykeln, en funktion som är specifik för den lilla isoformen. Första rapporten som föreslog JADE1-funktion i G2/M/G1-övergång visade att under den sena G2-fasen genomgår JADE1S fosforylering kopplad till dess dissociation från kromatin till cytoplasman. Massspektral analys identifierade att totalt sex individuella aminosyrarester fosforyleras av ett mitotiskt kinas. Baserat på farmakologisk analys regleras JADE1-fosforylering och kompartmentalisering av Aurora A- och Aurora B-vägar. Andra kinaser har rapporterats och kan spela en roll. Efter avslutad mitos kring telofas genomgår huvudpoolen av JADE1S-proteinet defosforylering och återassocieras med uppenbarligen kondenserande kromatin inuti de reformerade kärnorna. En diskret pool av JADE1S associeras med klyvningsfåran och uppträder därefter i mitten av den cytokinetiska bryggan. Endast JADE1S, men inte JADE1L eller HBO1 hittades i mitten av cellerna som genomgick cytokines. Den rumsliga regleringen av JADE1S under celldelningen föreslog roll i G2/M till G1-övergång, vilket inkluderar cytokines och slutlig abscission. Cytokinesis är det sista steget i cellcykeln som kontrollerar uppdelningen av cellinnehåll, inklusive cytoplasma, membran och kromatin. Cytokinetisk brygga skärs av under den sista abcissionen som sker nära mittkroppen och kan ta upp till 2 timmar. Cytokinesis och slutlig abscission kontrolleras hårt av regulatoriska proteinkomplex och checkpointproteiner. Antalet rapporter om cytokineskontroll har ökat under det senaste decenniet.
JADE1 roll i cytokines demonstrerades genom användning av flera funktionella analyser och cellodlingsmodeller. DNA-profilering av FACS visade att JADE1S-utarmning underlättade hastigheten av G1-cellers ackumulering i synkront delande HeLa-celler. Utarmningen av JADE1S-protein i asynkront delande celler minskade andelen cytokinetiska celler och ökade andelen multinukleära celler. Data visade att JADE1 negativt kontrollerar cytokines, förmodligen genom att bidra till cytokinesfördröjning. JADE1-nedreglering ökade antalet multinukleära celler som tyder på misslyckad cytokines, medan JADE1S måttligt överuttryck ökade antalet cytokinetiska celler som tyder på cytokinetisk fördröjning. Hämning av Aurora B-kinas av specifika småmolekylära läkemedel resulterade i frisättning av JADE1S-medierad cytokinetisk fördröjning och tillät progression av abscission. Eftersom Aurora B är en nyckelregulator för NoCut, kommer JADE1S sannolikt att reglera cytokines vid abscissionskontrollpunktskontrollen. JADE1S men inte JADE1L eller HBO1 hittades i centrosomer av delande celler under hela cellcykeln, och inget av dessa proteiner hittades i cilia. Däremot rapporterade en annan studie JADE1-lokalisering till flimmerhåren och centrosomen. Studien kommunicerade inte om JADE1 isoformspecificitet. Centrosomer är cytoskelettets kärnbildningscentra. Centrosomsignalering bidrar till definitionen av cellform, motilitet, orientering, polaritet, delningsplan och till troheten hos systerkromosomseparationen under mitos och cytokines.
pVHL
Den första proteinpartnern till JADE1S identifierades 2002 i en studie som letade efter nya partners till pVHL, som är en tumörsuppressor. Ett fåtal uppföljningsstudier karakteriserade bindning och gav några insikter om funktionella interaktioner av JADE1-pVHL. Den mänskliga pVHL är muterad i von Hippel-Lindau ärftlig sjukdom och i majoriteten av sporadiska klarcelliga njurkarcinom. Egenskaper och funktion hos pVHL har undersökts i många decennier och omfattande litteratur finns tillgänglig. En av de mer kända funktionerna hos pVHL är att förmedla protein ubiquitination och proteosomal nedbrytning. Som en komponent i ubiquitinligas E3-komplexet binder pVHL och riktar sig mot flera kända faktorer, inklusive HIF1a och HIF2a för ubiquitinering. Mekanism för HIF1a-aktivering genom hypoxi och roll för pVHL i denna väg har rapporterats för över ett decennium sedan. VHL-proteinet har studerats intensivt och kopplingen mellan naturligt förekommande mutationer och cancer har fastställts. Andra orsakande HIF-1a-oberoende pVHL-vägar har övervägts. Den fysiska interaktionen pVHL-JADE1S identifierades genom jäst-två hybridscreeningsanalys och bekräftades ytterligare biokemiskt. Samtransfektion av pVHL ökade JADE1S-proteinets halveringstid och överflöd, vilket tyder på potentiellt positivt samband. Vissa pVHL-cancerhärledda trunkationer men inte punktmutationer minskade pVHL-JADE1-stabiliseringsfunktionen, vilket tyder på koppling till pVHL-associerade cancerformer. Molekylära vägar och cellulär betydelse av JADE1-pVHL-interaktioner är inte väl förstått. Enskild studie som beskriver JADE1S inneboende ubiquitin-ligasaktivitet och ubiquitination av beta-catenin har rapporterats år 2008. Baserat på den studien har en modell föreslagits att pVHL reglerar beta-catenin genom JADE1, och PHD-zinkfingrar krävs för denna aktivitet.
Apoptos
JADE1S funktion vid apoptos har föreslagits men mekanismerna förblir svårfångade och resultaten är svåra att förena. Enligt studier bromsar JADE1-överuttryck hastigheten av celltillväxt och inducerar cellcykelstoppprotein p21. Flera försök att etablera pålitliga cellinjer som stabilt uttrycker JADE1S-protein har inte varit framgångsrika, förmodligen på grund av den negativa cellselektionen. I motsats till det visar en annan studie att JADE1-nedreglering minskade hastigheten för DNA-syntes i synkront delande celler. Enligt indirekt immunfluorescens- och mikroskopianalys av odlade celler orsakar odlade celler överbelastning med JADE1-protein celltoxicitet och biverkningar. Celler genomgår morfologiska förändringar som inte liknar apoptos men tyder på allvarligt försämrad cellcykel inklusive färgning av celler med onormala former och stora, multilobulära kärnor. Baserat på JADE1S-medierad reglering av cellcykeln övervägs andra tolkningar: JADE1-överbelastning kan orsaka förlängd NoCut och avstannad cytokines eller allvarlig cellcykelfelbalans snarare än direkt transkriptionsaktivering av apoptos.
Biologisk roll
Den biologiska rollen för JADE1 har inte klarlagts. Begränsat antal publikationer tar upp denna fråga med hjälp av mössmodeller. Den mest omfattande studien som publicerades 2003, identifierade möss ortolog av mänsklig JADE1, Jade1, och undersökte Jade1 uttryck under möss embryogenes. När de letade efter utvecklingsreglerade gener använde författarna genfällningsanalys och identifierade mus Jade1 som en starkt reglerad gen under embryogenesen. Insättning av vektorn i den tredje intronen av Jade1-genen leder till produktionen av ett 47-aminosyror trunkerat protein. Genfällans insättningsmutation resulterade i Jade1-beta-galaktosidasreporterfusionsprodukt och Jade1 nollallel. Även om homozygoterna för integreringen av genfällan inte producerade någon stark utvecklingsfenotyp, avslöjade fusionsprodukten Jade1-genens rumsliga-temporala uttryck i musembryonceller och vävnader från utvecklande embryon upp till 15,5 dpc. Dessutom rapporterar studien experimentell och jämförande silicoanalys av Jade1-mRNA-transkript, Jade1-genstruktur och analys av Jade1-proteinortologer från mus- och zebrafiskar. Jade1-uttryck detekterades i extraembryonala ektoderm och trofoblaster, som är placentakomponenter som är viktiga för vaskulogenes, såväl som på platser berikade med multipotenta eller vävnadsspecifika progenitorer, inklusive neurala progenitorer(2). Dynamiken i Jade1-reporteruttrycket i dessa områden indikerar involveringen i bestämningen och förlängningen av den främre bakre axeln, en viktig punkt i studien). Den potentiella rollen för human JADE1 i förnyelsen av embryonala stamcells- och embryonala karcinomcellkulturer föreslogs i en annan screeningstudie som visade att aktivering av stamcellstranskriptionsfaktor OCT4-vägen uppreglerade JADE1-genuttrycket tillsammans med stamcellsfaktorerna NANOG i odlade stamceller . , PHC1, USP44 och SOX2. Rollen för JADE1 i epitelcellsproliferation togs upp i en murin modell av akut njurskada och regenerering. Uttrycksmönster och dynamik hos HBO1-JADE1S/L undersöktes i regenererande tubulära epitelceller. Ischemi och reperfusionsskada resulterade i en initial minskning av proteinnivåerna JADE1S, JADE1L och HBO1, som återgick till baslinjen under njuråterhämtning. Uttrycksnivåer av HBO1 och JADE1S återhämtade sig när cellproliferationshastigheten nådde maximum, medan JADE1L återhämtade sig efter att bulkproliferationen hade minskat. Det tidsmässiga uttrycket av JADE1 korrelerade med acetyleringen av histon H4 (H4K5 och H4K12) men inte det för histon H3 (H4K14), vilket tyder på att JADE1-HBO1-komplexet specifikt markerar H4 under epitelcellsproliferation. Resultaten av studien implicerar JADE1-HBO1-komplex vid akut njurskada och föreslår distinkta roller för JADE1-isoformer under epitelcellsåtervinning.
Sjukdomsföreningar
Rollen för JADE1 i mänskliga sjukdomar har inte klarlagts. En nyligen genomförd studie sökte efter nya submikroskopiska genetiska förändringar i myelofibros, som är en benmärgscancer. Studien identifierade sju nya deletioner och translokationer i en liten kohort av patienter med primär myelofibros. JADE1 och den intilliggande genen som kallas Sodium channel och clathrin linker 1 (SCLT1) modifierades signifikant. Som ett resultat av mutation har JADE1-genen deletioner av intron 5-6 och exon 6-11, vilket skulle producera JADE1 som saknar en stor bit av protein från PHD-zinkfingret. Relevansen för patogenes är under utredning. I en handfull pilotstudier undersöktes uttryck av JADE1 i tjocktarmscancer och njurkarcinom. Resultaten i dessa studier stämmer inte alltid överens. Resultaten av vissa studier genereras mestadels från den histokemiska analysen av tumörprover med användning av JADE1-antikroppar med okarakteriserade specificiteter mot JADE1 i allmänhet, och JADE1S eller JADE1L i synnerhet. Resultat av studier som använder in silico mikroarray algoritmanalys visar att PHF17 mRNA kan spela en roll i utvecklingen av cancer i bukspottkörteln. Dessa lovande utredningslinjer kräver ytterligare kontroller och ytterligare bedömningar.
Interaktioner
Flera proteiner interagerar med JADE1, inklusive: pVHL, TIP60, HBO1, ING4, ING5, β-catenin, NPHP4.
Anteckningar
Vidare läsning
- Tzouanacou E, Tweedie S, Wilson V (december 2003). "Identifiering av Jade1, en gen som kodar för ett PHD-zinkfingerprotein, i en genfälla-mutagenesskärm för gener involverade i utveckling av anteroposterior axel" . Molekylär och cellulär biologi . 23 (23): 8553–2. doi : 10.1128/MCB.23.23.8553-8562.2003 . PMC 262661 . PMID 14612400 .
- Panchenko MV, Zhou MI, Cohen HT (december 2004). "von Hippel-Lindau partner Jade-1 är en transkriptionell samaktivator associerad med histonacetyltransferasaktivitet" . Journal of Biological Chemistry . 279 (53): 56032–41. doi : 10.1074/jbc.M410487200 . PMID 15502158 .
- Zhou MI, Foy RL, Chitalia VC, Zhao J, Panchenko MV, Wang H, Cohen HT (augusti 2005). "Jade-1, en kandidat för njurtumörsuppressor som främjar apoptos" . Proceedings of the National Academy of Sciences of the United States of America . 102 (31): 11035–40. Bibcode : 2005PNAS..10211035Z . doi : 10.1073/pnas.0500757102 . PMC 1182408 . PMID 16046545 .
- Doyon Y, Cayrou C, Ullah M, Landry AJ, Côté V, Selleck W, Lane WS, Tan S, Yang XJ, Côté J (januari 2006). "ING-tumörsuppressorproteiner är kritiska regulatorer av kromatinacetylering som krävs för genomuttryck och vidmakthållande" . Molekylär cell . 21 (1): 51–64. doi : 10.1016/j.molcel.2005.12.007 . PMID 16387653 .
- Lim J, Hao T, Shaw C, Patel AJ, Szabó G, Rual JF, Fisk CJ, Li N, Smolyar A, Hill DE, Barabási AL, Vidal M, Zoghbi HY (maj 2006). "Ett protein-proteininteraktionsnätverk för mänskliga ärftliga ataxier och störningar av Purkinje-celldegeneration" . Cell . 125 (4): 801–14. doi : 10.1016/j.cell.2006.03.032 . PMID 16713569 . S2CID 13709685 .
- Olsen JV, Blagoev B, Gnad F, Macek B, Kumar C, Mortensen P, Mann M (november 2006). "Global, in vivo och platsspecifik fosforyleringsdynamik i signalnätverk" . Cell . 127 (3): 635–48. doi : 10.1016/j.cell.2006.09.026 . PMID 17081983 . S2CID 7827573 .
externa länkar
- PHF17+protein,+human vid US National Library of Medicine Medical Subject Headings (MeSH)
Denna artikel innehåller text från United States National Library of Medicine, som är allmän egendom .