Homo naledi
|
Homo naledi Tidsintervall: Mellan pleistocen
|
|
|---|---|

|
|
| De 737 kända elementen i H. naledi | |
| vetenskaplig klassificering | |
| Rike: | Animalia |
| Provins: | Chordata |
| Klass: | Mammalia |
| Beställa: | primater |
| Underordning: | Haplorhini |
| Infraordning: | Simiiformes |
| Familj: | Hominidae |
| Underfamilj: | Homininae |
| Stam: | Hominini |
| Släkte: | Homo |
| Arter: |
† H. naledi
|
| Binomialt namn | |
|
† Homo naledi
Berger et al ., 2015
|
|

|
|
| Placering av Rising Star Cave i the Cradle of Humankind , Sydafrika | |
Homo naledi är en utdöd art av arkaisk människa som upptäcktes 2013 i Rising Star Cave , Cradle of Humankind , Sydafrika som dateras till Mellan Pleistocen för 335 000–236 000 år sedan. Den första upptäckten omfattar 1 550 exemplar, representerande 737 olika grundämnen, och minst 15 olika individer. Trots detta exceptionellt höga antal exemplar är deras klassificering med andra Homo fortfarande oklar.
Tillsammans med likheter med samtida Homo , delar de flera egenskaper med urfaderns Australopithecus och tidiga Homo ( mosaikutveckling ), framför allt en liten kraniell kapacitet på 465–610 cm 3 (28,4–37,2 cu in), jämfört med 1 270–1 330 cm 3 (78–81 cu in) hos moderna människor. De beräknas ha varit i genomsnitt 143,6 cm (4 fot 9 tum) i höjd och 39,7 kg (88 lb) i vikt, vilket ger en liten encefaliseringskvot på 4,5. Icke desto mindre H. naledi hjärnanatomi ha liknat den samtida Homo , vilket skulle kunna indikera jämförbar kognitiv komplexitet. Uthålligheten hos småhjärniga människor så länge mitt bland samtiden med större hjärnor reviderar den tidigare uppfattningen att en större hjärna nödvändigtvis skulle leda till en evolutionär fördel, och deras mosaikanatomi utökar kraftigt det kända variationsområdet för släktet.
H. naledis anatomi indikerar att även om de var kapabla till långväga resor med ett mänskligt steg och gång, var de mer trädlevande än andra Homo , bättre anpassade till klättring och upphängningsbeteende i träd än uthållighetslöpning . Tandans anatomi tyder på konsumtion av grynig mat täckt av partiklar som damm eller smuts. någon indikation på materiell kultur , verkar de ha varit skickliga nog att producera och hantera verktyg och troligen tillverkade tidig- eller medelstenåldersindustrier . Det har också kontroversiellt postulerats att dessa individer fick begravningsriter och bars in i och placerades i kammaren. I december 2022 rapporterades förslag om att H. naledi använde eld för ljus och matlagning.
Upptäckt

Den 13 september 2013 när de utforskade Rising Star Cave- systemet i the Cradle of Humankind , Sydafrika, hittade grottmänniska Rick Hunter och Steven Tucker homininfossiler längst ner i Dinaledi-kammaren . Den 24:e återvände de till kammaren och tog foton, som de visade för de sydafrikanska paleoantropologerna Pedro Boshoff och Lee Rogers Berger den 1 oktober. Berger sammanställde därefter ett utgrävningsteam som inkluderade Hunter och Tucker, de så kallade " Underground Astronauts " .
Kammaren hade gått in minst en gång tidigare, av grottor i början av 1990-talet. De ordnade om några ben och kan ha orsakat ytterligare skador, även om mycket av golvet i kammaren inte hade trampats på före 2013. Det ligger cirka 80 m (260 fot) från huvudentrén, längst ner på en 12 m (260 fot) 39 fot) vertikalt fall, och den 10 m (33 fot) långa huvudpassagen är bara 25–50 cm (10 in – 1 fot 8 in) som smalast. Totalt har mer än 1 550 benbitar som tillhör minst femton individer (9 omogna och 6 vuxna) återvunnits från de lerrika sedimenten. Berger och kollegor publicerade resultaten 2015.
Fossilerna representerar 737 anatomiska element – inklusive delar av skallen, käken, revbenen, tänderna, lemmar och innerörat ben – från gamla, vuxna, unga och infantila individer. Det finns också några ledade eller nära-artikulerade element, inklusive skallen med käkbenet, och nästan kompletta händer och fötter. Med antalet individer av båda könen över flera åldersdemografier är det den rikaste samlingen av associerade fossila homininer som upptäckts i Afrika. Bortsett från Sima de los Huesos- samlingen och senare prover från neandertalare och moderna människor, har utgrävningsplatsen den mest omfattande representationen av skelettelement över hela livslängden och från flera individer i fossilregistret för hominin.
Holotypprovet , DH1, består av en manlig partiell calvaria (överst på skallen), partiell maxilla och nästan komplett käkben . Paratyperna , DH2 till DH5, omfattar alla partiell calvaria . Berger och kollegor namngav arten Homo naledi , artnamnet som betyder "stjärna" på Sotho-språket , eftersom resterna kom från Rising Star Cave.
Kvarlevorna av minst tre ytterligare individer (två vuxna och ett barn) rapporterades i Lesedi-kammaren i grottan av John Hawks och kollegor 2017.
Klassificering
År 2017 daterades Dinaledi-lämningarna till 335 000–236 000 år sedan i Mellersta Pleistocen , med hjälp av elektronspinresonans (ESR) och uran-thorium (U-Th) datering på tre tänder, och U-Th och paleomagnetisk datering av sedimenten de deponerades i. Fossilerna ansågs tidigare ha daterats till 1~2 miljoner år sedan eftersom inga liknande småhjärniga homininer tidigare hade varit kända från ett sådant nyligen datum i Afrika (den mindre-hjärniga Homo floresiensis i Indonesien levde på en isolerad ö, och uppenbarligen utrotades kort efter de moderna människornas ankomst.) Förmågan hos en sådan småhjärnig hominin att ha överlevt så länge mitt i den större hjärna Homo reviderar avsevärt tidigare föreställningar om mänsklig evolution och uppfattningen att en större hjärna skulle nödvändigtvis leda till en evolutionär fördel. Deras mosaikenatomi utökar också avsevärt variationsområdet för släktet.
H. naledi antas ha förgrenats mycket tidigt från samtida Homo . Det är oklart om de förgrenade sig vid tiden för H. habilis , H. rudolfensis och A. sediba ; är ett systertaxon till H. erectus och den samtidiga storhjärniga Homo ; eller är ett systertaxon till ättlingarna till H. heidelbergensis (moderna människor och neandertalare). Detta skulle innebära att de avgrenade sig från samtida Homo senast före 900 000 år sedan, och möjligen så tidigt som Pliocen . Det är också möjligt att deras förfäder artade sig efter en korsningshändelse mellan Homo och sena australopitheciner. Om man tittar på skallen, H. naledi den närmaste affiniteten till H. erectus .
Det är oklart om dessa H. naledi var en isolerad befolkning i mänsklighetens vagga, eller om de sträckte sig över Afrika. Om det senare, så kan flera gracila hominin-fossiler över hela Afrika som traditionellt har klassificerats som sena H. erectus potentiellt representera H. naledi- exemplar.

Anatomi
Skalle
.jpg)
Två manliga H. naledi- skallar från Dinaledi-kammaren hade kranialvolymer på cirka 560 cm 3 (34 cu in), och två honskallar 465 cm 3 (28,4 cu in). En manlig H. naledi- skalle från Lesedi-kammaren hade en kraniell volym på 610 cm 3 (37 cu in). Dinaledi-exemplaren liknar mer kranialkapaciteten hos australopithecines; Som jämförelse H. erectus i genomsnitt cirka 900 cm 3 (55 cu in), och moderna människor 1 270 och 1 130 cm 3 (78 och 69 cu in) för män respektive honor. Lesedi-exemplaret ligger mer inom intervallet för H. habilis och H. e. georgicus . Icke desto mindre encefaliseringskvoten för H. naledi till 3,75, vilket är samma som pygmén H. floresiensis , men anmärkningsvärt mindre än alla andra Homo . Samtida Homo var alla över 6, H. e. georgicus på 3,55, och A. africanus på 3,81. Det är oklart om H. naledi ärvde liten hjärnstorlek från den senaste gemensamma Homo- förfadern, eller om den utvecklades sekundärt på senare tid.
Skalleformen är mer lik Homo , med en smalare form, närvaron av tinning- och nacklober i hjärnan och minskad postorbital sammandragning (skallen blir inte smalare bakom ögonhålorna). Pannlobsmorfologin är mer eller mindre densamma i alla Homo- hjärnor trots storlek, och skiljer sig från Australopithecus , som har varit inblandad i framställning av verktyg, utveckling av språk och socialitet.
Liksom moderna människor, men till skillnad från fossila homininer (inklusive sydafrikanska australopithecines, H. erectus och neandertalare), bröt den permanenta 2:a molaren ut relativt sent i livet, och dök upp tillsammans med premolarerna istället för tidigare, vilket indikerar en långsammare mognad som är ovanligt jämförbar med moderna människor . Framtändernas tandbildningshastighet är också mest lik moderna människor. Den övergripande storleken och formen på molarerna liknar mest de av tre oidentifierade Homo- exemplar från de lokala Swartkrans- och östafrikanska Koobi Fora -grottorna, och liknar i storlek (men inte form) Pleistocene H. sapiens . Halsarna på kindtänderna är proportionellt lika de hos A. afarensis och Paranthropus . Till skillnad från moderna människor och samtida Homo , saknar H. naledi flera accessoriska tandegenskaper och har en hög frekvens av individer som uppvisar huvudknutar, nämligen metakonen ( mittlinjen på tungans sida) och hypokonen (till höger på läppsidan). ) på 2:a och 3:e molarerna, och en Y-formad hypoconulid (en ås på läppsidan mot kinden) på alla 3 molarerna. Icke desto mindre H. naledi också många dentala likheter med samtida Homo .
Städet (ett mellanörat ben ) liknar mer de hos schimpanser, gorillor och Paranthropus än Homo . Liksom H. habilis och H. erectus har H. naledi en välutvecklad ögonbrynsrygg med en spricka som sträcker sig över strax ovanför åsen, och liksom H. erectus en uttalad nackknöl . H. naledi har vissa ansiktslikheter med H. rudolfensis .
Bygga

H. naledi -exemplaren uppskattas i genomsnitt ha stått runt 143,6 cm (4 fot 9 tum) och vägt 39,7 kg (88 lb). Denna kroppsmassa är mellanliggande mellan vad som vanligtvis ses hos Australopithecus och Homo -arter. Liksom andra Homo var hanen och honan H. naledi troligen ungefär lika stora, hanarna i genomsnitt cirka 20 % större än honorna. Ett ungt exemplar, DH7, överensstämmer skelettmässigt med en tillväxthastighet som liknar de snabbare apaliknande banorna för MH1 ( A. sediba ) och Turkana boy ( H. ergaster ). Men eftersom tandutvecklingen är så lik den moderna människors, är en långsammare mognadshastighet inte helt utesluten. Med den snabbare tillväxttakten skulle DH7 ha dött vid 8–11 år gammal, men med den långsammare tillväxten skulle DH7 ha dött vid 11–15 år gammal.
Beträffande ryggraden är endast den 10:e och 11:e bröstkotan (i bröstregionen) bevarade från förmodligen en enda individ, som är proportionellt lika de av samtida Homo , fastän är de minsta registrerade av någon hominin. Kotans två tvärgående processer, som sticker ut diagonalt, liknar mest neandertalarnas. Neurala kanalerna inom är proportionellt stora, liknar moderna människor, neandertalare och H. e. georgicus . Det 11:e revbenet är rakt som det hos A. afarensis , och det 12:e revbenet är robust i tvärsnitt som neandertalarnas. Liksom neandertalarna verkar det 12:e revbenet ha stöttat starka interkostala muskler ovanför, och en stark quadratus lumborum-muskel under. Men till skillnad från neandertalarna var det svagt fäste vid diafragman . Sammantaget verkar detta H. naledi -exemplar ha varit liten kropp jämfört med andra Homo , även om det är oklart om detta enstaka exemplar är representativt för arten.
Skuldrorna är mer lika de hos australopithecines, med skulderbladet placerat högre på ryggen och längre från mittlinjen, korta nyckelben och liten eller ingen humerus torsion. Förhöjda axel- och nyckelben indikerar en smal bröstkorg. Bäckenet och benen har särdrag som påminner om Australopithecus , inklusive anterposteriort sammanpressade (framifrån till baksida) lårbenshackar , mediolateralt sammanpressade (från vänster till höger) tibiae och en något cirkulär fibulär hals ; vilket tyder på en bred buk. Denna kombination skulle utesluta effektiv uthållighetslöpning hos H. naledi , till skillnad från H. erectus och ättlingar. Istället H. naledi ha varit mer trädlevande .
Lemmer


Det metakarpala benet i tummen var välutvecklat, vilket används för att hålla och manipulera stora föremål, och hade starka toppar för att stödja dess opponens pollicis-muskel som används vid precisions-nypa grepp, och dess sedanmuskler . Detta är mer likt andra Homo än Australopithecus . H. naledi verkar ha haft starka flexor pollicis longus-muskler som moderna människor, med människoliknande handflats- och fingerkuddar, som är viktiga för ett kraftfullt grepp mellan tummen och fingrarna. Men till skillnad från Homo är H. naledi tummetakarpalleden jämförbart liten i förhållande till tummens längd, och tummens phalangeal- leden är tillplattad. Det distala tumfalanxbenet är robust och proportionellt mer likt de hos H. habilis och P. robustus .
De andra fingrarnas metacarpals delar anpassningar med moderna människor och neandertalare för att kupa och manipulera föremål, och handledsleden är överlag jämförbar med den hos moderna människor och neandertalare. Omvänt är de proximala falangerna krökta och är nästan identiska med de hos A. afarensis och H. habilis , vilket tolkas som en anpassning för klättring och upphängningsbeteende . Sådan krökning är mer uttalad hos vuxna än ungdomar, vilket tyder på att vuxna klättrade lika mycket eller mer än ungdomar, och detta beteende gjordes ofta. Fingrarna är också proportionellt längre än de hos någon annan fossil hominin (förutom den trädlevande Ardipithecus ramidus och ett modernt mänskligt exemplar från Qafzeh-grottan, Israel) vilket också överensstämmer med klättringsbeteende.
H. naledi var tvåbent och stod upprätt. Liksom andra Homo hade de stark insättning för sätesmusklerna , väldefinierade linea aspera (en ås som löper ner på baksidan av lårbenet), tjocka patellae , långa tibiae och gracila fibulae. Dessa indikerar att de var kapabla till långväga resor. H. naledi -foten liknade den hos moderna människor och andra Homo , med anpassningar för bipedalism och en mänsklig gång. Hälbenet har dock en låg orientering, jämförbar med den hos icke-mänskliga människoapor, och ankelbenet har en låg deklination, vilket möjligen indikerar att foten skulle ha varit subtilt styvare under ställningsfasen av promenaden innan foten tryckte av marken.
Patologi
Den vuxna högra underkäken UW 101-1142 har en benig lesion, vilket tyder på en godartad tumör . Individen skulle ha upplevt viss svullnad och lokalt obehag, men tumörens position nära den mediala pterygoidmuskeln (som sannolikt orsakar obehag på käkens gångjärn) kan ha hämmat muskelns funktion och förändrat höjden av höger sida av käken.
Dentala defekter i H. naledi- prover under 1,6–2,8 och 4,3–7,6 månaders utveckling orsakades med största sannolikhet av säsongsbetonade stressorer. Detta kan ha berott på extrema sommar- och vintertemperaturer som orsakar matbrist. Dessutom är lägsta vintertemperaturer i området i genomsnitt cirka 3 °C (37 °F), och kan sjunka under fryspunkten; H. naledi med liten kropp, och vintrarna ökade sannolikt mottagligheten för luftvägssjukdomar. Miljöpåfrestningar stämmer överens med att dagens influensasäsonger i Sydafrika når sin topp under vintern, och att pediatrisk diarré sjukhusvård är vanligast vid höjden av regnperioden på sommaren.
Lokala homininer var sannolikt rovdjur av stora köttätare, såsom lejon, leoparder och hyenor. Det verkar dock finnas en tydlig brist på stora rovdjursrester från den norra änden av mänsklighetens vagga, där Rising Star Cave ligger, möjligen för att köttätare föredrog Blaaubank-floden söderut som kan ha erbjudit bättre jaktmarker med en större överflöd av stora bytesobjekt. Alternativt, eftersom många fler platser är kända i söder än i norr, kanske rovdjurens rumsliga mönster inte är väl representerade av fossilregistret ( bevarande bias) .
Kultur

Mat
Tandspån och slitage indikerar den vanliga konsumtionen av små hårda föremål, såsom smuts och damm, och koppformad slitage på de bakre tänderna kan ha härrört från grusiga partiklar. Dessa kan ha härrört från otvättade rötter och knölar. Alternativt kan torrhet ha rört upp partiklar på matvaror och belagt maten med damm. Det är möjligt att de vanligtvis åt större hårda föremål, såsom frön och nötter, men dessa bearbetades till mindre bitar före konsumtion. Hur som helst, H. naledi ockuperade en till synes unik nisch från tidigare sydafrikanska homininer, inklusive Australopithecus och Paranthropus . Tänderna hos alla tre arterna indikerar dock att de behövde utöva hög skjuvkraft för att tugga igenom kanske växt- eller muskelfibrer. Tänderna hos andra Homo kan inte producera så höga krafter, kanske på grund av användningen av vissa livsmedelsbearbetningstekniker, såsom matlagning. I december 2022 rapporterades förslag om att H. naledi använde eld för ljus och matlagning.
Teknologi
Även om H. naledi- lämningar inte är förknippade med några stenverktyg, är det troligt att de producerade tidig stenålder ( Acheulean och möjligen den tidigare Oldawan ) eller medelstenåldersindustrin eftersom de har samma anpassningar till handen som andra mänskliga arter som är inblandade i verktygstillverkning. H. naledi är den enda identifierade mänskliga arten som har funnits under den tidiga medelstenåldern i Highveld -regionen, Sydafrika, vilket möjligen indikerar att denna art tillverkade och upprätthöll denna tradition åtminstone under denna tidsperiod. Sådana industrier och stenskärningstekniker har troligen utvecklats oberoende flera gånger bland olika Homo- arter och -populationer, eller transporterades över långa avstånd av uppfinnarna eller lärlingarna och undervisades.
Begravningar

År 2015 drog arkeologen Paul Dirks, Berger och kollegor slutsatsen att kropparna medvetet måste ha burits och placerats in i kammaren av människor eftersom de verkar ha varit intakta när de först deponerades i kammaren (inga bevis på trauma genom att vara släpps in i kammaren eller av predation, och exceptionell bevarande), kammaren är otillgänglig för stora rovdjur, kammaren verkar vara ett isolerat system och har aldrig översvämmats (det vill säga naturkrafter var inte i spel), det finns inget gömt axel genom vilken människor av misstag kunde ha ramlat in genom, och det finns inga bevis på någon katastrof som dödade alla individer inne i kammaren. De sa att det också är möjligt att kropparna tappades ner i en ränna och föll långsamt på grund av oregelbundenhet och trånghet på vägen ner, eller en mjuk lerkudde att landa på. I båda scenarierna skulle morticians ha krävt artificiellt ljus för att navigera grottan; och platsen användes upprepade gånger för begravningar eftersom kropparna inte alla deponerades samtidigt.
2016 kontrade paleoantropologen Aurore Val att sådan bevarande kan ha berott på mumifiering snarare än noggrann begravning, och frånvaron av långa benhuvuden påminner om predation, och hon anser att det är orättfärdigt att bortse från naturkrafter som översvämningar för att deponera kropparna. Vidare finns det bevis på skador orsakade av skalbaggar, skalbaggarlarver och sniglar (vilket underlättar nedbrytning); men kammaren uppvisar inte idealiska förhållanden för sniglar, och den innehåller inte heller snigelskal, vilket skulle tyda på att nedbrytning faktiskt initierades före avsättningen i kammaren.
Icke desto mindre bekräftade Dirks, Berger och kollegor 2017 att det inte finns några bevis på att vatten rinner in i grottan, och att det är mer troligt att dessa H. naledi begravdes i kammaren. De sa också att det är möjligt att de begravdes av samtida Homo , såsom förfäderna till moderna människor, snarare än andra H. naledi , men inte desto mindre att det kulturella beteendet för begravningsmetoder inte är omöjligt för H. naledi och begravning. i kammaren kan ha gjorts för att ta bort ruttnande kroppar från en bosättning, förhindra asätare eller på grund av social bindning och sorg.
2018 upprepade antropologen Charles Egeland och kollegor Vals känslor och konstaterade att det inte finns tillräckligt med bevis för att dra slutsatsen att mänskliga arter hade utvecklat ett koncept om livet efter detta så tidigt i tiden. De sa också att bevarandet av Dinaledi-individerna liknar de av babiankadaver som samlas i grottor (antingen genom naturlig död av grottlevande babianer eller genom att en leopard släpar in kadaver).
År 2021, efter analysen av benfragmenten från en omogen individ, hävdade Juliet Brophy och Berger återigen "Vi antar att kranialresterna och tänderna som vi har samlat in på UW 110 har resulterat från in situ fragmentering av en partiell eller fullständig kranium deponerades på denna avlägsna plats. Vi ser det som troligt att någon homininbyrå var inblandad i deponeringen av kranialmaterialet."
Galleri

(A,B) Digitalt rekonstruerade skallsidor
Underkäkarna på LES1 (vänster) och DH1 (höger)
Övre käkar på LES1 (vänster) och DH1 (höger)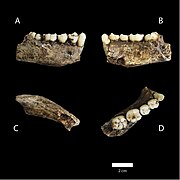 (A,B,C,D) Vy över ena underkäken
(A,B,C,D) Vy över ena underkäken Utsikt över en nyckelbenet
Utsikt över en nyckelbenet Utsikt över ena humerus
Utsikt över ena humerus Utsikt över en ulna
Utsikt över en ulna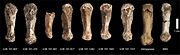 Metacarpals från flera individer, var och en märkt
Metacarpals från flera individer, var och en märkt Utsikt över en 10: e bålkota
Utsikt över en 10: e bålkota Utsikt över en 11: e bålkota
Utsikt över en 11: e bålkota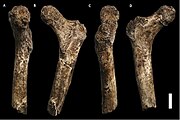 Utsikt över ena lårbenet
Utsikt över ena lårbenet (A,B,C,D) Vy över ett skenben
(A,B,C,D) Vy över ett skenben Ankelben från flera individer, var och en märkt
Ankelben från flera individer, var och en märkt (1) vuxen höger fot, (2) juvenil vänster, (3,4) vuxen vänster, (5) juvenil höger
(1) vuxen höger fot, (2) juvenil vänster, (3,4) vuxen vänster, (5) juvenil höger















Se även
- Afrikansk arkeologi
- Australopithecus sediba – Två miljoner år gammal hominin från mänsklighetens vagga
- Denisovan – asiatisk arkaisk människa
- Homo luzonensis – arkaisk människa från Luzon, Filippinerna
- Homo floresiensis – arkaisk människa från Flores, Indonesien
- Neanderthal – Utdöda eurasiska arter eller underarter av arkaiska människor
- Tidslinje för mänsklig evolution
Vidare läsning
- Berger, LR ; Hawks, JD (2017). Nästan mänsklig: Den häpnadsväckande berättelsen om Homo naledi och upptäckten som förändrade vår mänskliga berättelse . Washington, DC: National Geographic Society . ISBN 978-1-4262-1811-8 .
externa länkar
- Rekonstruktioner av H. naledi av paleoartist John Gurche
- Wheeler, Sharon. "Sändningar från en av grottornas Rising Stars" . Mörker nedanför .
- "Prominenta hominidfossiler" . Talk Origins .
- "Utforska den hominida fossila rekorden" . Bradshaw Foundation .
- "blogg för medlemmar i Rising Star Expedition" . National Geographic . Arkiverad från originalet den 9 september 2015.
- "Tredimensionella skanningar av Homo naledi- fossiler" . MorphoSource .
- "Human Timeline (Interactive)" . Naturhistoriska riksmuseet . Smithsonian .
|
Taxonomi ( Homininer ) |
|||||||||
|---|---|---|---|---|---|---|---|---|---|
| Förfäder |
|
||||||||
| Modeller |
|
||||||||
| Tidslinjer | |||||||||
| Andra | |||||||||
| Webbplatser |  |
||||
|---|---|---|---|---|---|
| Fossiler | |||||
| Antropologi |
|
||||
| Myndighetskontroll : Nationella bibliotek |
|---|