Orbitofrontal cortex
| Orbitofrontal cortex | |
|---|---|
 Ungefärlig placering av OFC visas på en sagittal MRI
| |
 Orbital yta av vänster frontallob.
| |
| Detaljer | |
| Del av | Frontallob |
| Identifierare | |
| latin | cortex orbitofrontalis |
| Neuronamn | 91 |
| NeuroLex ID | birnlex_1049 |
| FMA | 242003 |
| Anatomiska termer för neuroanatomi | |
Orbitofrontal cortex ( OFC ) är en prefrontal cortexregion i hjärnans frontallober som är involverad i den kognitiva processen för beslutsfattande . Hos icke-mänskliga primater består den av associationsbarkområdena Brodmann-område 11 , 12 och 13 ; hos människor består den av Brodmann område 10 , 11 och 47 .
OFC är funktionellt relaterat till den ventromediala prefrontala cortexen . Därför särskiljs regionen på grund av de distinkta neurala anslutningarna och de distinkta funktioner den utför. Det definieras som den del av den prefrontala cortexen som tar emot projektioner från den mediala dorsala kärnan i thalamus , och tros representera känslor , smak, lukt och belöning i beslutsfattande. Den har fått sitt namn från sin position omedelbart ovanför de banor där ögonen befinner sig. Betydande individuell variation har hittats i OFC hos människor. Ett relaterat område finns hos gnagare .
Strukturera
OFC är uppdelad i flera breda regioner som kännetecknas av cytoarkitektur, inklusive brodmann område 47/12 , brodmann område 11 , brodmann område 14 , brodmann område 13 och brodmann område 10 . Fyra gyri delas av ett komplex av sulci som oftast liknar ett "H"- eller ett "K"-mönster. Sträcker sig längs den rostro-caudala axeln, två sulci, de laterala och orbitala sulci, är vanligtvis förbundna med den transversella orbital sulcus, som sträcker sig längs en medial-lateral axel. Mest medialt är den mediala orbital gyrus separerad från gyrus rectus av olfactory sulcus. Anteriort består både gyrus rectus och den mediala delen av mediala orbital gyrus av area 11(m), och posteriort, område 14. Den posteriora orbital gyrus består till största delen av område 13, och avgränsas medialt och lateralt av de främre extremiteterna på de mediala och laterala orbitala sulci. Område 11 utgör en stor del av OFC som involverar både de laterala delarna av den mediala orbitala gyrusen såväl som den främre orbitala gyrusen. Den laterala orbitala gyrusen består till största delen av området 47/12. Det mesta av OFC är granulärt , även om de kaudala delarna av område 13 och område 14 är agranulära. Dessa kaudala regioner, som ibland inkluderar delar av den insulära cortexen , reagerar främst på obearbetade sensoriska signaler.
Anslutningar
OFC:s anslutningsmöjligheter varierar något längs en rostral-caudal axel. Den kaudala OFC är tyngre sammankopplad med sensoriska regioner, särskilt tar emot direkt input från den pyriforma cortexen . Den kaudala OFC är också den som är mest sammankopplad med amygdala . Rostralt tar OFC emot färre direkta sensoriska projektioner och är mindre ansluten till amygdala, men den är sammankopplad med den laterala prefrontala cortex och parahippocampus . OFC:s anslutningsmöjligheter har också konceptualiserats som att den består av två nätverk; ett orbitalnätverk som består av de flesta av de centrala delarna av OFC, inklusive de flesta av områdena 47/12, 13 och 11; ett medialt nätverk som består av de mediala mest och caudolaterala regionerna av OFC, såväl som områdena 24 , 25 och 32 av den mediala prefrontala cortex. De mediala och orbitala nätverken kallas ibland för det "visceromotoriska nätverket" respektive det "sensoriska nätverket".
Afferenter
OFC tar emot projektioner från flera sensoriska modaliteter. Den primära olfaktoriska cortex , gustatoriska cortex , sekundära somatosensoriska cortex , superior och inferior temporal gyrus (som förmedlar visuell information) projekterar alla till OFC. Evidensen för auditiv input är svag, även om vissa neuroner reagerar på auditiva stimuli, vilket indikerar att en indirekt projektion kan förekomma. OFC tar också emot input från den mediala dorsala kärnan , insulär cortex , entorhinal cortex , perirhinal cortex , hypotalamus och amygdala .
Efferenter
Den orbitofrontala cortex är ömsesidigt förbunden med perirhinal och entorhinal cortex, amygdala, hypotalamus och delar av den mediala temporalloben. Utöver dessa utgångar projicerar OFC också till striatum, inklusive nucleus accumbens , caudate nucleus och ventral putamen , såväl som regioner av mellanhjärnan inklusive det periaqueductal gråa och ventrala tegmentala området . OFC-ingångar till amygdalasynapsen på flera mål, inklusive två robusta vägar till basolaterala amygdala och interkalerade celler i amygdala, samt en svagare direkt projektion till amygdalas centrala kärna .
Fungera
Flera funktioner har tillskrivits OFC, inklusive att förmedla kontextspecifikt svar, kodning av oförutsedda händelser på ett flexibelt sätt, kodningsvärde, kodning av antaget värde, inhiberande svar, inlärning av oförutsedda förändringar, känslomässig bedömning, ändra beteende genom somatiska markörer, drivande socialt beteende och representerar tillståndsutrymmen. Medan de flesta av dessa teorier förklarar vissa aspekter av elektrofysiologiska observationer och lesionsrelaterade förändringar i beteende, misslyckas de ofta med att förklara, eller motsägs av andra fynd. Ett förslag som förklarar mångfalden av OFC-funktioner är att OFC kodar tillståndsutrymmen, eller den diskreta konfigurationen av interna och externa egenskaper associerade med en situation och dess oförutsedda händelser. Till exempel kan förslaget att OFC kodar ekonomiskt värde vara en återspegling av Tillståndsvärde för OFC-kodningsuppgift. Representationen av uppgiftstillstånd skulle också kunna förklara förslaget att OFC fungerar som en flexibel karta över oförutsedda händelser, eftersom ett byte av uppgiftstillstånd skulle möjliggöra kodning av nya oförutsedda händelser i ett tillstånd, med bevarande av gamla oförutsedda händelser i ett separat tillstånd, vilket möjliggör byte av oförutsedda händelser när det gamla uppgiftstillståndet blir relevant igen. Representationen av uppgiftstillstånd stöds av elektrofysiologiska bevis som visar att OFC svarar på en mångfald av uppgiftsfunktioner och kan snabbt kartläggas under oförutsedda skift. Representationen av uppgiftstillstånd kan påverka beteendet genom flera potentiella mekanismer. Till exempel är OFC nödvändigt för i ventralt tegmentalt område (VTA) för att producera ett dopaminergt belöningsförutsägelsefel, och OFC kan koda förväntningar för beräkning av RPE i VTA.
Specifika funktioner har tillskrivits underregioner av OFC. Den laterala OFC har föreslagits för att återspegla potentiellt valvärde, vilket möjliggör fiktiva (kontrafaktiska) prediktionsfel för att potentiellt förmedla växlingsval under reversering, utplåning och devalvering. Optogenetisk aktivering av lOFC förbättrar mål riktat över vanemässigt beteende, vilket möjligen återspeglar ökad känslighet för potentiella val och därför ökad byte. mOFC, å andra sidan, har föreslagits för att återspegla relativt subjektivt värde. Hos gnagare har en liknande funktion tillskrivits mOFC, kodande aktionsvärde på ett graderat sätt, medan lOFC har föreslagits för att koda specifika sensoriska egenskaper hos resultaten. LOFC har också föreslagits för att koda associationer för stimulusresultat, som sedan jämförs med värde i mOFC. Metaanalys av neuroimagingstudier hos människor avslöjar att det finns en medial-lateral valensgradient, där den mediala OFC svarar oftast på belöning och den laterala OFC svarar oftast på straff. En posterior-främre abstrakthetsgradient hittades också, där den bakre OFC svarade på enklare belöning och den främre OFC svarade mer på abstrakta belöningar. Liknande resultat rapporterades i en metaanalys av studier om primära kontra sekundära belöningar.
OFC och basolateral amygdala (BLA) är starkt sammankopplade, och deras anslutning är nödvändig för devalveringsuppgifter. Skador på antingen BLA eller OFC före, men endast OFC efter devalvering försämrar prestandan. Medan BLA endast svarar på signaler som förutsäger framträdande utfall på ett graderat sätt i enlighet med värde, reagerar OFC på både värde och de specifika sensoriska attributen hos cue-outcome associationer. Medan OFC-neuroner som, tidigt i inlärningen, reagerar på resultatmottagning, normalt överför sitt svar till uppkomsten av signaler som förutsäger resultatet, försämrar skador på BLA denna form av inlärning.
Den bakre orbitofrontala cortex (pOFC) är ansluten till amygdala via flera vägar, som kan både uppreglera och nedreglera det autonoma nervsystemets aktivitet. Preliminära bevis tyder på att neuromodulatorn dopamin spelar en roll i att förmedla balansen mellan de hämmande och excitatoriska vägarna, med ett högt dopamintillstånd som driver autonom aktivitet.
Det har föreslagits att den mediala OFC är involverad i att skapa stimulans-belöningsassociationer och med förstärkning av beteende, medan den laterala OFC är involverad i stimulus-outcome associationer och utvärdering och eventuellt vändning av beteende. Aktivitet i den laterala OFC finns till exempel när försökspersoner kodar för nya förväntningar om straff och sociala repressalier.
OFC i mitten av främre delen har visat sig konsekvent spåra subjektivt nöje i neuroavbildningsstudier. En hedonisk hotspot har upptäckts i den främre OFC, som kan förbättra tyckesvaret på sackaros. OFC kan också påverka de affektiva svaren som induceras av α-amino-3-hydroxi-5-metyl-4-isoxazolpropionsyra (AMPA) -antagonism i kärnan hopar sig mot aptitliga svar.
OFC är kapabel att modulera aggressivt beteende via projektioner till interneuroner i amygdala som hämmar glutaminerga projektioner till den ventromediala hypotalamus .
Elektrofysiologi
Neuroner i OFC svarar både på primära förstärkare, såväl som signaler som förutsäger belöningar över flera sensoriska domäner. Bevisen för svar på visuella, smakmässiga, somatosensoriska och luktstimuli är robusta, men bevisen för hörselsvar är svagare. I en delmängd av OFC-neuroner, moduleras neurala svar på belöningar eller belöningssignaler av individuella preferenser och av interna motivationstillstånd som hunger. En bråkdel av neuroner som reagerar på sensoriska signaler som förutsäger en belöning är selektiva för belöning och uppvisar ett vändningsbeteende när signalutfallsrelationer byts ut. Neuroner i OFC uppvisar också svar på frånvaron av en förväntad belöning och bestraffning. En annan population av neuroner uppvisar svar på nya stimuli och kan "komma ihåg" välbekanta stimuli i upp till en dag.
Under cued belöning eller cued instrumental belöning uppgifter, neuroner i OFC uppvisar tre allmänna mönster av skjutning; skjuta som svar på signaler; skjuta före belöning mottagande; skjuta som svar på belöningsmottagandet. I motsats till den mediala prefrontala cortex och striatum uppvisar OFC-neuroner inte avfyring som förmedlar genom rörelse. Deras belöningsförutsägande svar formas dock av uppmärksamhet: när man flyttar uppmärksamheten mellan två alternativ kommer samma OFC-population att representera positivt värdet av ett föremål som för närvarande besöks, men negativt värdet på det obevakade föremålet. Kodningen av belöningsstorleken är också flexibel och tar hänsyn till de relativa värdena för nuvarande belöningar.
Människor
Den mänskliga OFC är bland de minst förstådda regionerna i den mänskliga hjärnan. Det har föreslagits att OFC är involverat i sensorisk integration, i att representera det affektiva värdet av förstärkare och i beslutsfattande och förväntningar. Särskilt OFC verkar vara viktigt för att signalera förväntade belöningar/straff för en handling med tanke på de särskilda detaljerna i en situation. Genom att göra detta kan hjärnan jämföra den förväntade belöningen/straffet med den faktiska leveransen av belöning/straff, vilket gör OFC avgörande för adaptivt lärande. Detta stöds av forskning på människor, icke-mänskliga primater och gnagare.
Psykiatriska störningar
Den orbitofrontala cortexen har varit inblandad i borderline personlighetsstörning , schizofreni , egentlig depression , bipolär sjukdom , tvångssyndrom , missbruk , posttraumatisk stressyndrom , autism och panikångest . Även om neuroimagingstudier har gett bevis för dysfunktion i en mängd olika psykiatriska störningar, komplicerar den gåtfulla karaktären av OFC:s roll i beteendet förståelsen av dess roll i patofysiologin för psykiatriska störningar. Funktionen av OFC är inte känd, men dess anatomiska kopplingar med ventral striatum, amygdala, hypotalamus, hippocampus och periaqueductal grått stödjer en roll i att förmedla belönings- och rädslarelaterade beteenden.
Tvångssyndrom
Metaanalyser av neuroimagingstudier vid OCD rapporterar hyperaktivitet i områden som allmänt anses vara en del av det orbitofrontala segmentet av den kortiko-basala ganglia-thalamo-kortikala slingan såsom caudate nucleus , thalamus och orbitofrontal cortex. OCD har föreslagits för att återspegla en positiv återkopplingsslinga på grund av ömsesidig excitation av OFC och subkortikala strukturer. Medan OFC vanligtvis är överaktiv under symtomprovokerande uppgifter, framkallar kognitiva uppgifter vanligtvis hypoaktivitet hos OFC; detta kan återspegla en distinktion mellan känslomässiga och icke-emotionella uppgifter, laterala och mediala OFC, eller helt enkelt bara inkonsekventa metoder.
Missbruk
Djurmodeller och cellspecifika manipulationer i förhållande till drogsökande beteende implicerar dysfunktion hos OFC vid missbruk. Missbruksstörningar är förknippade med en mängd olika brister relaterade till flexibelt målinriktat beteende och beslutsfattande. Dessa brister överlappar med symptom relaterade till OFC-lesioner och är också associerade med minskad orbitofrontal grå substans , hypometabolism i vilotillstånd och trubbig OFC-aktivitet under uppgifter som involverar beslutsfattande eller målinriktat beteende. I motsats till vilotillstånd och beslutsrelaterad aktivitet framkallar signaler associerade med droger robust OFC-aktivitet som korrelerar med begär. Gnagarstudier visar också att lOFC till BLA-projektioner är nödvändiga för cue-inducerad återinsättning av självadministration. Dessa fynd är alla kongruenta med den roll som OFC spelar för att koda resultaten förknippade med vissa stimuli. Utvecklingen mot tvångsmässigt missbruk kan återspegla ett skifte mellan modellbaserat beslutsfattande, där en intern modell för framtida resultat vägleder beslut, till modell för fritt lärande, där beslut baseras på förstärkningshistoria. Modellbaserat lärande involverar OFC och är flexibelt och målinriktat, medan modellfritt lärande är mer rigid; eftersom ett övergång till mer modellfritt beteende på grund av dysfunktion i OFC, som det som produceras av missbruk av droger, kan ligga till grund för drogsökningsvanor.
Beteendestörningar
Uppförandestörning är förknippad med både strukturella abnormiteter och funktionella abnormiteter under affektiva uppgifter. Avvikelser i OFC-struktur, aktivitet och funktionell anslutning har alla observerats i samband med aggression.
Affektiva störningar
Neuroimagingstudier har funnit abnormiteter i OFC vid MDD och bipolär sjukdom. I överensstämmelse med den mediala/belönings- och laterala/straffgradienten som finns i neuroimagingstudier, har vissa neuroimagingstudier observerat förhöjd lateral OFC-aktivitet vid depression, såväl som minskad sammankoppling av den mediala OFC, och förbättrad sammankoppling i den laterala OFC. Hypoaktivitet av lateral OFC har ofta observerats vid bipolär sjukdom, särskilt under maniska episoder.
Forskning
Avbildning
Att använda funktionell magnetisk resonanstomografi (fMRI) för att avbilda den mänskliga OFC är en utmaning, eftersom denna hjärnregion ligger i närheten av de luftfyllda bihålorna . Detta innebär att artefaktfel kan uppstå i signalbehandlingen , vilket till exempel orsakar geometriska förvrängningar som är vanliga när man använder ekoplanär avbildning (EPI) vid högre magnetfältstyrkor. Extra försiktighet rekommenderas därför för att få en bra signal från den orbitofrontala cortex, och ett antal strategier har utarbetats, såsom automatisk shimsning vid höga statiska magnetfältstyrkor.
Gnagare
Hos gnagare är OFC helt agranulär eller dysgranulär. OFC är indelat i ventrolaterala (VLO), laterala (LO), mediala (MO) och dorsolaterala (DLO) regioner. Genom att använda mycket specifika tekniker för att manipulera kretsar, såsom optogenetik , har OFC varit inblandad i OCD-liknande beteenden och i förmågan att använda latenta variabler i beslutsfattande uppgifter.
Klinisk signifikans
Skada
Förstörelse av OFC genom förvärvad hjärnskada leder vanligtvis till ett mönster av ohämmat beteende. Exempel inkluderar överdrivet svordomar, hypersexualitet, dålig social interaktion, spelberoende, droganvändning (inklusive alkohol och tobak) och dålig empatisk förmåga. Hämmat beteende hos patienter med vissa former av frontotemporal demens tros vara orsakat av degeneration av OFC.
Avbrott
När OFC-förbindelser störs kan ett antal kognitiva, beteendemässiga och emotionella konsekvenser uppstå. Forskning stöder att de huvudsakliga störningarna som är förknippade med oreglerad OFC-anslutning/kretslopp kretsar kring beslutsfattande, känsloreglering, impulsiv kontroll och belöningsförväntningar. En nyligen genomförd multimodal human neuroimaging studie visar störd strukturell och funktionell anslutning av OFC med de subkortikala limbiska strukturerna (t.ex. amygdala eller hippocampus) och andra frontala regioner (t.ex. dorsal prefrontal cortex eller främre cingulate cortex) korrelerar med onormal OFC-påverkan ( t.ex. rädsla) bearbetning hos kliniskt oroliga vuxna.
En tydlig förlängning av problem med beslutsfattande är drogberoende/ substansberoende , som kan bli följden av störningar i striato-thalamo-orbitofrontala kretsen. Attention Deficit Hyperactivity Disorder (ADHD) har också varit inblandad i dysfunktion av neurala belöningskretsar som kontrollerar motivation, belöning och impulsivitet, inklusive OFC-system. Andra störningar av exekutiv funktion och impulskontroll kan påverkas av OFC-kretsdysreglering, såsom tvångssyndrom och trikotillomani
Vissa demenssjukdomar är också förknippade med störningar i OFC-anslutningen. Beteendevarianten av frontotemporal demens är associerad med neurala atrofimönster av projektionsfibrer av vit och grå substans involverade i OFC-anslutning. Slutligen tyder en del forskning på att senare stadier av Alzheimers sjukdom påverkas av förändrad anslutning av OFC-system.
Orbitofrontal epilepsi
Orbitofrontal epilepsi är sällsynt, men förekommer. Presentationen av OFC-epilepsi är ganska varierande, även om vanliga egenskaper inkluderar sömnrelaterade symtom, automatismer och hypermotoriska symtom. En recension rapporterade att auror i allmänhet inte var vanliga eller ospecifika, medan en annan rapporterade att OFC-epilepsi var associerad auror som involverade somatosensoriska fenomen och rädsla.
bedömning
Visuell diskrimineringstest
Det visuella diskrimineringstestet har två komponenter. I den första komponenten, "reversal learning", presenteras deltagarna för en av två bilder, A och B. De lär sig att de kommer att belönas om de trycker på en knapp när bild A visas, men straffas om de trycker på knappen när bild. B visas. När denna regel har fastställts byter regeln. Med andra ord, nu är det korrekt att trycka på knappen för bild B, inte bild A. De flesta friska deltagare uppfattar denna regelomvändning nästan omedelbart, men patienter med OFC-skada fortsätter att svara på det ursprungliga mönstret av förstärkning, även om de är nu straffas för att ha hållit ut med det. Rolls et al. noterade att detta beteendemönster är särskilt ovanligt med tanke på att patienterna rapporterade att de förstod regeln.
Den andra komponenten i testet är "utrotning". Återigen lär sig deltagarna att trycka på knappen för bild A men inte bild B. Men den här gången, istället för att reglerna ändras, ändras regeln helt. Nu kommer deltagaren att straffas för att ha tryckt på knappen som svar på endera bilden. Det korrekta svaret är att inte trycka på knappen alls, men personer med OFC-dysfunktion har svårt att motstå frestelsen att trycka på knappen trots att de blir straffade för det.
Iowa speluppgift
Iowa- speluppgiften är en simulering av beslutsfattande i verkligheten och används ofta i kognition och känsloforskning. Deltagarna presenteras med fyra virtuella kortlekar på en datorskärm. De får veta att varje gång de väljer ett kort står de för att vinna lite spelpengar. De får höra att målet med spelet är att vinna så mycket pengar som möjligt. Men då och då kommer de att förlora lite pengar när de väljer ett kort. Uppgiften är tänkt att vara ogenomskinlig, det vill säga att deltagarna inte är avsedda att medvetet utarbeta regeln, och de är tänkta att välja kort baserat på sin "magreaktion " . Två av däcken är "dåliga däck", vilket betyder att de under tillräckligt lång tid kommer att göra en nettoförlust; de andra två kortlekarna är "bra kortlekar" och kommer att göra en nettovinst över tiden.
De flesta friska deltagare provar kort från varje kortlek, och efter cirka 40 eller 50 val är de ganska bra på att hålla sig till de bra kortlekarna. Patienter med OFC-dysfunktion fortsätter dock att hålla ut med de dåliga kortlekarna, ibland även om de vet att de förlorar pengar totalt sett. Samtidig mätning av galvanisk hudrespons visar att friska deltagare visar en "stress"-reaktion på att sväva över de dåliga däcken efter bara 10 försök, långt innan de medvetna känner att däcken är dåliga. Däremot utvecklar patienter med OFC-dysfunktion aldrig denna fysiologiska reaktion på förestående straff. Bechara och hans kollegor förklarar detta i termer av den somatiska markörhypotesen . Iowa-speluppgiften används för närvarande av ett antal forskargrupper som använder fMRI för att undersöka vilka hjärnregioner som aktiveras av uppgiften hos friska frivilliga såväl som kliniska grupper med tillstånd som schizofreni och tvångssyndrom .
Faux pas-testet är en serie vinjetter som berättar om ett socialt tillfälle då någon sa något som inte borde ha sagts, eller en obekväm händelse. Deltagarens uppgift är att identifiera vad som sades som var besvärligt, varför det var besvärligt, hur människor skulle ha känt sig som reaktion på faux pas och på en saklig kontrollfråga. Även om testet först utformades för användning hos personer inom autismspektrumet , är testet också känsligt för patienter med OFC-dysfunktion, som inte kan bedöma när något socialt besvärligt har hänt trots att de verkar förstå historien mycket väl.
Se även
Ytterligare bilder
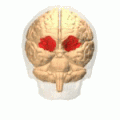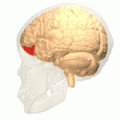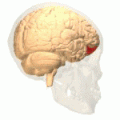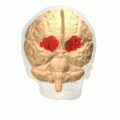
Orbital gyrus visas i rött.

Mediala ytan av hjärnbarken - gyri

Storhjärnans basala yta. Orbital gyrus visas i rött.

Lateral orbitofrontal cortex

Medial orbitofrontal cortex, inre skivavy.
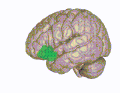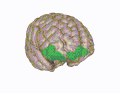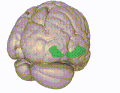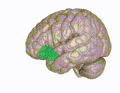
3D-visualisering av den orbitofrontala cortex i en genomsnittlig mänsklig hjärna

Orbitofrontal cortex markerad i grönt på koronala T1 MRI-bilder

Orbitofrontal cortex markerad i grönt på sagittal T1 MRI-bilder

Orbitofrontal cortex markerad i grönt på transversala T1 MRI-bilder









externa länkar
- Cerebral Cortex specialutgåva om orbitofrontal cortex
- Camille et al. (2004) Den orbitofrontala cortexens inblandning i upplevelsen av ånger