Opabinia
|
Opabinia Tidsintervall: Mellankambrium ( Wuliuan ),
|
|
|---|---|

|
|
| Fossilt exemplar ( lektotyp ) visas på Smithsonian i Washington, DC, | |
| vetenskaplig klassificering | |
| Rike: | Animalia |
| Provins: | Arthropoda |
| Klass: | † Dinocaridida |
| Familj: | † Opabiniidae |
| Släkte: |
† Opabinia Walcott , 1912 |
| Arter: |
† O. regalis
|
| Binomialt namn | |
|
† Opabinia regalis
Walcott , 1912
|
|
Opabinia regalis är en utdöd , stamgruppsleddjur som finns i den mellankambriska Burgess Shale Lagerstätte (505 miljoner år sedan) i British Columbia . Opabinia var ett mjukt djur, som mätte upp till 7 cm i kroppslängd, och dess segmenterade bål hade flikar längs sidorna och en solfjäderformad svans. Huvudet visar ovanliga drag: fem ögon , en mun under huvudet och vänd bakåt, och en kload snabel som troligen förde mat till munnen. Opabinia levde förmodligen på havsbotten och använde snabeln för att leta efter små, mjuka föda. Färre än tjugo bra exemplar har beskrivits; 3 exemplar av Opabinia är kända från Greater Phyllopod bed , där de utgör mindre än 0,1% av samhället.
När den första grundliga undersökningen av Opabinia 1975 avslöjade dess ovanliga särdrag, troddes den inte vara relaterad till någon känd filum , eller kanske en släkting till leddjurs- och annelidförfäder . Senare studier sedan slutet av 1990-talet stöder dock konsekvent dess affinitet som en medlem av basala leddjur, tillsammans med de nära besläktade radiodonts ( Anomalocaris och släktingar) och gällobopodier ( Kerygmachela och Pambdelurion ).
På 1970-talet pågick en debatt om huruvida flercelliga djur dök upp plötsligt under det tidiga kambrium, i en händelse som kallas den kambriska explosionen , eller hade uppstått tidigare men utan att lämna fossiler. Till en början betraktades Opabinia som starka bevis för den "explosiva" hypotesen. Senare upptäckten av en hel serie liknande lobopodiska djur, några med närmare likheter med leddjur, och utvecklingen av idén om stamgrupper antydde att det tidiga kambrium var en tid med relativt snabb evolution men en tid som kunde förstås utan att anta någon unik evolutionära processer.
Upptäcktshistoria
År 1911 hittade Charles Doolittle Walcott i Burgess Shale nio nästan kompletta fossil av Opabinia regalis och några av vad han klassificerade som Opabinia? media och publicerade en beskrivning av alla dessa 1912. Det generiska namnet kommer från Opabin-passet mellan Mount Hungabee och Mount Biddle , sydost om Lake O'Hara , British Columbia, Kanada . 1966–1967 Harry B. Whittington ett annat bra exemplar, och 1975 publicerade han en detaljerad beskrivning baserad på mycket noggrann dissektion av några exemplar och fotografier av dessa exemplar upplysta från en mängd olika vinklar. Whittingtons analys täckte inte Opabinia? media : Walcotts exemplar av denna art kunde inte identifieras i hans samling. 1960 beskrev ryska paleontologer exemplar som de hittade i Norilsky-regionen i Sibirien och märkte Opabinia norilica, men dessa fossiler var dåligt bevarade, och Whittington ansåg inte att de gav tillräckligt med information för att klassificeras som medlemmar av släktet Opabinia .
Förekomst
Alla de erkända Opabinia- exemplaren som hittills hittats kommer från " Phyllopod bed " i Burgess Shale, i de kanadensiska klippiga bergen i British Columbia.
1997 rapporterade Briggs och Nedin från södra Australien Emu Bay Shale ett nytt exemplar av Myoscolex som var mycket bättre bevarat än tidigare exemplar, vilket fick dem att dra slutsatsen att det var en nära släkting till Opabinia - även om denna tolkning senare ifrågasattes av Dzik, som drog istället slutsatsen att Myoscolex var en annelidmask .
Morfologi
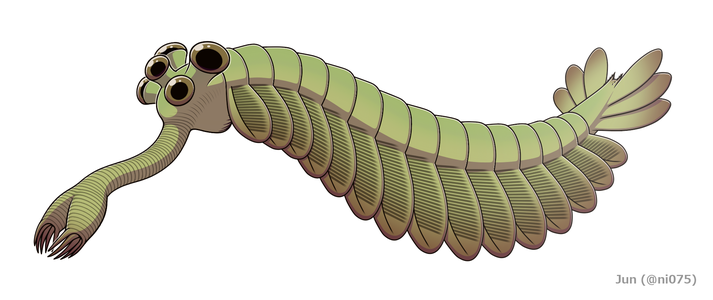
Restaurering

Storleksuppskattning



Opabinia såg så märklig ut att publiken vid den första presentationen av Whittingtons analys skrattade. Längden på Opabinia regalis från huvudet (exklusive snabel) till svansänden varierade mellan 4 centimeter (1,6 tum) och 7 centimeter (2,8 tum). En av de mest utmärkande karaktärerna hos Opabinia är den ihåliga snabeln , vars totala längd var ungefär en tredjedel av kroppens och projicerade ner från under huvudet. Snabeln var tvärstrimmig som en dammsugarslang och flexibel, och den slutade med en kloliknande struktur vars ändkanter bar 5 ryggar som sträckte sig inåt och framåt. Den bilaterala symmetrin och laterala (istället för vertikala som rekonstruerats av Whittington 1975) arrangemanget av klon antyder att det representerar ett par sammansmälta frontala bihang, jämförbara med de för radiodonts och gälade lobopodier . Huvudet bar fem skaftade ögon: två nära framsidan och ganska nära mitten av huvudet, pekade uppåt och framåt; två större ögon med längre stjälkar nära huvudets bakre och yttre kanter, pekande uppåt och åt sidan; och ett enda öga mellan det större paret av stjälkade ögon, som pekar uppåt. Det har antagits att ögonen alla var sammansatta , som andra leddjurs laterala ögon, men denna rekonstruktion, som inte backas upp av några bevis, är "något fantasifull". Munnen låg under huvudet, bakom snabeln och pekade bakåt , så att matsmältningskanalen bildade en U-böj på sin väg mot baksidan av djuret. Snabeln verkade tillräckligt lång och flexibel för att nå munnen.
Huvuddelen av kroppen var typiskt cirka 5 millimeter (0,20 tum) bred och hade 15 segment, på var och en av dem fanns par av flikar (lober) som pekade nedåt och utåt. Flikarna överlappade varandra så att framsidan av var och en täcktes av bakkanten på den framför sig. Kroppen slutade med vad som såg ut som ett enda koniskt segment med tre par överlappande svansfläktblad som pekade uppåt och utåt och bildade en svans som en V-formad dubbelfläkt.

![Various interpretations on the flap and gill structures of Opabinia regalis A: Whittington (1975),[2] B: Bergström (1986),[17] C: Budd (1996),[5] D: Zhang & Briggs (2007),[7] E: Budd & Daley (2011)[8]](//upload.wikimedia.org/wikipedia/commons/thumb/3/34/20210809_Opabinia_regalis_flap_gill_interpretation.png/780px-20210809_Opabinia_regalis_flap_gill_interpretation.png)
Olika tolkningar av flik- och gälstrukturerna hos Opabinia regalis A: Whittington (1975), B: Bergström (1986), C: Budd (1996), D: Zhang & Briggs (2007), E: Budd & Daley (2011)![Opabinia cross-section based on Budd and Daley (2011)[8]](//upload.wikimedia.org/wikipedia/commons/thumb/3/30/20210807_Opabinia_regalis_trunk_cross_section.png/420px-20210807_Opabinia_regalis_trunk_cross_section.png)
Opabinia -tvärsnitt baserat på Budd och Daley (2011)
![Various interpretations on the flap and gill structures of Opabinia regalis A: Whittington (1975),[2] B: Bergström (1986),[17] C: Budd (1996),[5] D: Zhang & Briggs (2007),[7] E: Budd & Daley (2011)[8]](https://commons.wikimedia.org/wiki/File:20210809_Opabinia_regalis_flap_gill_interpretation.png)
![Opabinia cross-section based on Budd and Daley (2011)[8]](https://commons.wikimedia.org/wiki/File:20210807_Opabinia_regalis_trunk_cross_section.png)
Tolkningar av andra egenskaper hos Opabinia -fossiler skiljer sig åt. Eftersom djuren inte hade mineraliserad rustning eller ens tuffa organiska exoskelett som hos andra leddjur, plattades deras kroppar till när de begravdes och fossiliserades, och mindre eller inre drag uppträder som markeringar inom konturerna av fossilerna.
Whittington (1975) tolkade gälarna som parade förlängningar fästa dorsalt till baserna på alla utom de första flikarna på varje sida, och trodde att dessa gälar var platta undertill, hade överlappande lager ovanpå. Bergström (1986) avslöjade att de "överlappande skikten" var rader av individuella blad, tolkade flikarna som en del av ryggbeläggningar ( tergite ) över kroppens övre yta, med blad fästa under var och en av dem. Budd (1996) trodde att gälbladen var fästa längs framkanterna på ryggsidan av alla utom de första flikarna. Han hittade också märken innanför flikarnas framkanter som han tolkade som inre kanaler som förbinder gälarna med det inre av kroppen, ungefär som Whittington tolkade märket längs snabeln som en inre kanal. Zhang och Briggs (2007) tolkade dock att alla flikar har ett bakre avstånd där gälbladen fästes. Budd och Daley (2011) förkastar rekonstruktionen av Zhang & Briggs, och visar att flikarna har kompletta posteroir-kanter som tidigare rekonstruktioner. De följer mestadels rekonstruktionen av Budd (1996) med modifieringar av vissa detaljer (t.ex. det första flikparet har också gälar; fästpunkten för gälbladen ligger dock mer baktill än tidigare).


Whittington (1975) hittade bevis på nästan triangulära drag längs kroppen och drog slutsatsen att de var inre strukturer, troligen sidledes förlängningar av tarmen ( diverticula ). Chen et al. (1994) tolkade dem som inneslutna i loberna längs sidorna. Budd (1996) tyckte att "trianglarna" var för breda för att passa in i Opabinias smala kropp, och att tvärsnittsvyer visade att de var fästa separat från och lägre än loberna och sträckte sig under kroppen . Han hittade senare exemplar som verkade bevara benens yttre nagelband. Han tolkade därför "trianglarna" som korta, köttiga, koniska ben (lobopoder). Han hittade även små mineraliserade fläckar i spetsarna på några och tolkade dessa som klor. Under denna rekonstruktion homologiserades den gälbärande klaffen och lobopoden till den yttre gälgrenen och den inre grenen av leddjurs biramösa lemmar som ses i Marrella , trilobiter och kräftdjur . Zhang och Briggs (2007) analyserade den kemiska sammansättningen av "trianglarna", och kom fram till att de hade samma sammansättning som tarmen, och höll därför med Whittington om att de var en del av matsmältningssystemet. Istället betraktade de Opabinias lob+gälarrangemang som en tidig form av leddjurens lemmar innan den splittrades till en biramös struktur . Denna liknande kemiska sammansättning är dock inte bara förknippad med matsmältningskanalen; Budd och Daley (2011) föreslår att det representerar mineralisering som bildas i vätskefyllda håligheter i kroppen, vilket överensstämmer med ihåliga lobopoder som ses i otvetydiga lobopodiska fossiler. De klargör också att tarmdivertiklarna hos Opabinia är serier av cirkulära tarmkörtlar individualiserade från "trianglarna". Även om de var överens om frånvaron av terminala klor, kvarstår förekomsten av lobopoder i Opabinia som en rimlig tolkning.
Livsstil
Det sätt som Burgess Shale- djuren begravdes på, av ett lerskred eller en sedimentladdad ström som fungerade som en sandstorm, tyder på att de levde på havsbottens yta. Opabinia använde troligen sin snabel för att söka i sedimentet efter matpartiklar och föra dem till munnen. Eftersom det inte finns några tecken på något som kan fungera som käkar, var dess mat förmodligen liten och mjuk. De parade tarmdivertiklarna kan öka effektiviteten i matsmältningen och intaget av näring. Whittington (1975) som trodde att Opabinia inte hade några ben, trodde att den kröp på sina lober och att den också kunde ha simmat långsamt genom att flaxa med loberna, speciellt om den tidsinställda rörelserna för att skapa en våg med lobernas metakorala rörelse. Å andra sidan tyckte han att kroppen inte var tillräckligt flexibel för att tillåta fiskliknande vågor av hela kroppen.
Klassificering
|
||||||||||||||||||||||||||||||||||||||||||
| Sammanfattad fylogeni mellan Opabinia och andra ecdysozoiska taxa. |






Med tanke på hur paleontologernas rekonstruktioner av Opabinia skiljer sig är det inte förvånande att djurets klassificering var mycket omdiskuterad under 1900-talet. Charles Doolittle Walcott , den ursprungliga beskrivaren , ansåg att det var ett anostracan kräftdjur 1912. Idén följdes av G. Evelyn Hutchinson 1930, vilket gav den första rekonstruktionen av Opabinia som en anostracan som simmade upp och ner. Alberto Simonetta tillhandahöll en ny rekonstruktion av Opabinia 1970 som skiljer sig mycket från Hutchinsons, med massor av leddjursfunktioner (t.ex. dorsala exoskelett och ledade lemmar) som påminner om Yohoia och Leanchoilia . Leif Størmer , efter tidigare arbete av Percy Raymond , trodde att Opabinia tillhörde de så kallade "trilobitoider" ( trilobiter och liknande taxa). Efter sin grundliga analys Harry B. Whittington slutsatsen att Opabinia inte var leddjur 1975, eftersom han inte hittade några bevis för leddjursledslemmar, och ingenting liknande den flexibla, förmodligen vätskefyllda snabeln var känd hos leddjur. Även om han lämnade Opabinias klassificering över familjenivån öppen, övertalade den annullerade men inte ledade kroppen och de ovanliga sidoflikarna med gälar honom att det kan ha varit en representant för förfädernas stam från ursprunget av annelider och leddjur, två distinkta djur phyla ( Lophotrochozoan respektive Ecdysozoan ) som fortfarande troddes vara nära släktingar (förenade under Articulata ) vid den tiden.
1985 publicerade Derek Briggs och Whittington en stor ombeskrivning av Anomalocaris , också från Burgess Shale. Strax efter det konstaterade svensk paleontolog Jan Bergström 1986 likheten mellan Anomalocaris och Opabinia , och föreslog att de två djuren var släkt, eftersom de delade många drag (t.ex. sidoflikar, gälblad, skaftade ögon och specialiserade frontala bihang). Han klassificerade dem som primitiva leddjur, även om han ansåg att leddjur inte är en enda filum .
1996 fann Graham Budd vad han ansåg bevis på korta ben utan leder i Opabinia . Hans undersökning av den gälade lobopoden Kerygmachela från Sirius Passet lagerstätte , ungefär och över 10 miljoner år äldre än Burgess-skiffern, övertygade honom om att detta exemplar hade liknande ben. Han ansåg att benen på dessa två släkten mycket liknar dem hos Burgess Shale lobopodier Aysheaia och de moderna onychophorans (sammetsmaskar), som anses bära många förfädersdrag som delas av förfäder med leddjur. Efter att ha undersökt flera uppsättningar egenskaper som delas av dessa och liknande lobopodier, gjorde han en "bredskalig rekonstruktion av leddjursstamgruppen", med andra ord av leddjur och vad han ansåg vara deras evolutionära basala medlemmar. En slående egenskap hos detta släktträd är att moderna tardigrader (vattenbjörnar) kan vara Opabinias närmaste evolutionära släktingar. Å andra sidan, Hou et al. (1995, 2006) föreslog snarare att Opabinia är en medlem av ovanliga cykloneurala maskar med konvergenta leddjursdrag.
Även om Zhang och Briggs (2007) inte höll med Budds diagnos att Opabinias "trianglar" var ben, ledde likheten de såg mellan Opabinias lob+gälarrangemang och leddjurs biramösa lemmar att de drog slutsatsen att Opabinia var mycket nära besläktad med leddjur . Faktum är att de presenterade ett släktträd mycket likt Budds förutom att deras inte nämnde tardigrader. Oavsett de olika morfologiska tolkningarna, drog alla större omstudier sedan 1980-talet på liknande sätt slutsatsen att likheten mellan Opabinia och leddjur (t.ex. stjälkade ögon; dorsal segmentering; posterior mun; sammansmälta bihang; gälliknande lemgrenar) är taxonomiskt signifikanta.
Sedan 2010-talet stöds inte längre den föreslagna nära relationen mellan Opabinia och tardigrades/cycloneuralians, medan affiniteten hos Opabinia som stamgruppsleddjur tillsammans med Radiodonta (en kladd som inkluderar Anomalocaris och dess släktingar) och gälade lobopodier är allmänt accepterad, konsekvent återhämtad genom multipel fylogenetisk analys, såväl som nya upptäckter såsom närvaron av leddjursliknande tarmkörtlar och mellanliggande taxon Kylinxia .
År 2022 beskrev paleontologer ett liknande djur som upptäcktes i kambriska åldrade klippor i Utah . Fossilen fick namnet Utaurora comosa . och hittades inom Wheeler Shale Stam-leddjuret beskrevs faktiskt först 2008, men vid den tiden ansågs det ursprungligen vara ett exemplar av Anomalocaris. Denna uppenbarelse kan tyda på att det fanns andra djur som såg ut som Opabinia, och dess familj kan ha varit mer mångsidig.
Teoretisk betydelse

Opabinia gjorde det klart hur lite som var känt om mjuka djur, som vanligtvis inte lämnar fossiler. När Whittington beskrev det i mitten av 1970-talet fanns det redan en livlig debatt om djurens tidiga evolution . Preston Cloud hävdade 1948 och 1968 att processen var "explosiv", och i början av 1970-talet utvecklade Niles Eldredge och Stephen Jay Gould sin teori om punkterad jämvikt , som ser evolution som långa intervall av nära-stasis "avbrutna" av korta perioder av snabb förändring. Å andra sidan, ungefär samtidigt hävdade Wyatt Durham och Martin Glaessner båda att djurriket hade en lång proterozoisk historia som gömdes av bristen på fossiler. Whittington (1975) drog slutsatsen att Opabinia och andra taxa som Marrella och Yohoia inte kan rymmas i moderna grupper. Detta var en av de främsta anledningarna till att Gould i sin bok om Burgess Shale , Wonderful Life , ansåg att det tidiga kambriska livet var mycket mer mångsidigt och "experimentellt" än någon senare uppsättning djur och att den kambriska explosionen var en verkligt dramatisk händelse, möjligen driven av ovanliga evolutionära mekanismer. Han ansåg Opabinia som så viktig för att förstå detta fenomen att han ville kalla sin bok Homage to Opabinia .
Men snart följde andra upptäckter och analyser, som avslöjade liknande djur som Anomalocaris från Burgess Shale och Kerygmachela från Sirius Passet . Ett annat Burgess Shale-djur, Aysheaia , ansågs mycket likt modern Onychophora , som betraktas som nära släktingar till leddjur. Paleontologer definierade en grupp som kallas lobopodier för att inkludera fossila panarthropoder som tros vara nära släktingar till onychophorans, tardigrades och arthropoder men saknar ledben. Denna grupp blev senare allmänt accepterad som en parafyletisk grad som ledde till uppkomsten av bevarade panarthropod phyla.

- — = Nedstigningslinjer
- = Basal nod
- = Kronnod
- = Total grupp
- = Krongrupp
- = Stamgrupp
Medan denna diskussion om specifika fossiler som Opabinia och Anomalocaris pågick i slutet av 20-talet, introducerades begreppet stamgrupper för att täcka evolutionära "tanter" och "kusiner". En krongrupp är en grupp närbesläktade levande djur plus deras sista gemensamma förfader plus alla dess avkomlingar. En stamgrupp innehåller utlöpare från medlemmar av släktlinjen tidigare än den sista gemensamma förfadern till krongruppen; det är ett relativt begrepp, till exempel tardigrader är levande djur som bildar en krongrupp i sin egen rätt, men Budd (1996) betraktade dem också som en stamgrupp i förhållande till leddjuren. Att se konstiga organismer som Opabinia på detta sätt gör det möjligt att se att även om den kambriska explosionen var ovanlig, kan den förstås i termer av normala evolutionära processer.
Se även
- Kroppsplan – Uppsättning av morfologiska egenskaper som är gemensamma för medlemmar av en djurgrupp
- Radiodonta – Utdöd ordning av kambriska leddjur
Vidare läsning
- Bergström, J. (1987). "Den kambriska Opabinia och Anomalocaris ". Lethaia . 20 (2): 187–188. doi : 10.1111/j.1502-3931.1987.tb02037.x .
- Briggs, DEG; Whittington, HB (1987). "Släktskapen mellan de kambriska djuren Anomalocaris och Opabinia ". Lethaia . 20 (2): 185–186. doi : 10.1111/j.1502-3931.1987.tb02036.x .
externa länkar
- " Opabinia regalis " . Burgess Shale Fossil Gallery . Kanadas virtuella museum. 2011. Arkiverad från originalet 2020-11-12.
- Smithsonian sida på Opabinia , med foto av Burgess Shale fossil