Extrakromosomalt cirkulärt DNA
Extrakromosomalt cirkulärt DNA ( eccDNA ) är en typ av dubbelsträngad cirkulär DNA-struktur som först upptäcktes 1964 av Alix Bassel och Yasuo Hotta. I motsats till tidigare identifierade cirkulära DNA-strukturer (t.ex. bakterieplasmider , mitokondrie -DNA , cirkulära bakteriella kromosomer eller kloroplast-DNA ), är eccDNA cirkulärt DNA som finns i de eukaryota kärnorna i växt- och djurceller (inklusive mänskliga). Extrakromosomalt cirkulärt DNA härrör från kromosomalt DNA, kan variera i storlek från 50 baspar till flera megabaspar i längd, och kan koda för regulatoriska element och fullängdsgener . eccDNA har observerats i olika eukaryota arter och det föreslås vara en biprodukt av programmerade DNA- rekombinationshändelser , såsom V(D)J-rekombination .
Historisk bakgrund
1964 publicerade Bassel och Hotta sin första upptäckt av eccDNA som de gjorde när de undersökte Franklin Stahls kromosomala teori. I sina experiment visualiserade de isolerade vetekärnor och galtspermier med hjälp av elektronmikroskopi . Deras forskning fann att galtspermier innehöll eccDNA av olika storlekar. År 1965 identifierade Arthur Spriggs forskargrupp eccDNA i proverna av fem pediatriska patienters embryonala tumörer och en vuxen patients bronkialkarcinom . Under de följande åren ledde ytterligare forskning till upptäckten av eccDNA i olika arter som anges i Tabell 1:
| År | Organism | Referens |
|---|---|---|
| 1965 | Galtspermier _ | Hotta och Bassel, 1965 |
| 1965 | Mänskliga tumörer | Cox et al., 1965 |
| 1969 | Jäst | Billheimer och Avers, 1969 |
| 1984 | Trypanosomatider | Beverly et al., 1984 |
| 1972 | Euglena | Nass och Ben-Shaul, 1972 |
| 1972 | Tobak | Wong och Wildman, 1972 |
| 1972, 1978, 1980 | Svampar | Agsteribbe et al., 1972; Stahl et al., 1978; Lazarus et al., 1980 |
| 1972, 1985 | Odlade mänskliga fibroblaster | Smith och Vinograd, 1972; Riabowol et al., 1985 |
| 1976 | Xenopus | Buongiorno-Nardelli et al., 1976 |
| 1978, 1984 | Kycklingbursa | DeLap och Rush, 1978; Toda och Yamagishi, 1984 |
| 1982 | Mänskliga vävnader | Calabretta et al., 1982 |
| 1983 | Mus embryo | Yamagishi et al., 1983 |
| 1983, 1988, 1990 | Musvävnader | Tsuda et al., 1983; Flores et al., 1988; Gaubatz och Flores, 1990 |
| 1983 | Mus tymocyter | Yamagishi et al., 1983 |
| 1983 | Muslymfocyter | Tsuda et al., 1983 |
2000-talets forskning
Under 2000-talet har forskare fokuserat på att bättre karakterisera de specifika subtyperna av eccDNA, såväl som strukturen och funktionen hos dessa molekyler inom biologiska system:
- År 2012, Shibata et al. upptäckte en specifik typ av eccDNA som kallas mikroDNA . Forskarna hittade tiotusentals mikroDNA i musvävnader och cellinjer, såväl som mänskliga cellinjer.
- År 2017, Turner et al. identifierat med hjälp av helgenomsekvensering (WGS), cytogenetisk analys och strukturell modellering att extrakromosomalt cirkulärt DNA är mycket amplifierat och vanligt i olika typer av cancer . De fann att eccDNA-molekyler har betydande heterogenitet mellan olika celler även om de härrör från samma individ. Dessutom innehöll dessa eccDNA-molekyler tumördrivande gener och rapporterades sällan hittas i icke-cancerösa vävnader.
- År 2018 har Møller et al. använde friska mänskliga muskel- och blodcellsprover för att identifiera över 100 000 typer av eccDNA, vilket antydde att eccDNA kunde hittas i somatiska celler överallt.
- År 2019, Wu et al. fann att ecDNA (undertyp av eccDNA) associerar med kromatin , men till skillnad från kromosomer har det inte högre ordningskomprimering, vilket ökar dess tillgänglighet.
- År 2021, Wang et al. utvecklade bildandet av eccDNA och identifierade den immunstimulerande funktionen hos eccDNA. De utvecklade också ett förbättrat eccDNA-reningsprotokoll som minskar linjär DNA-kontamination i renade prover.
eccDNA-rening
Historiskt sett renades eccDNA med hjälp av en tvåstegsprocedur som involverade först isolering av rå extrakromosomalt DNA och därefter smältning av linjärt DNA via exonukleas - digestion . Ändå resulterar denna teknik ofta i linjär DNA-kontamination eftersom exonukleas-digestion inte är tillräcklig för att avlägsna allt linjärt DNA. År 2021, Wang et al. utvecklat en trestegs eccDNA-berikningsmetod som förbättrade eccDNA-rening:
- Cellerna dehydrerades först i > 90 % metanol. För att extrahera rå extrakromosomalt DNA lyserades cellerna med en alkalisk lysbuffert med pH 11,8, neutraliserades med en neutraliseringsbuffert och fälldes ut med användning av en utfällningsbuffert. En kommersiell plasmidreningskits silikakolonn användes för att isolera DNA från andra cellkomponenter.
- Det eluerade DNA:t digererades med restriktionsenzymet Pacl för att linjärisera mitokondrie-DNA ( mtDNA ) och ett exonukleas som kan smälta linjärt DNA.
- Slutligen utvanns cirkulärt DNA selektivt av en kommersiell lösning och kiseldioxidpärlor för att avlägsna linjärt DNA som inte avlägsnades genom exonukleas-digestion.
Dubbla minuter (DM) kontra extrakromosomalt cirkulärt DNA (eccDNA)
användes termen dubbelminuter (DM) ofta för att hänvisa till extrakromosomalt cirkulärt DNA eftersom det ofta förekom som ett par i tidiga studier. Allt eftersom forskningen har fortsatt har olika subtyper av extrakromosomalt cirkulärt DNA identifierats som inte är dubbla minuter (t.ex. mikroDNA ) . År 2014, Barreto et al. identifierade att dubbla minuter endast omfattar ungefär 30% av extrakromosomalt DNA. Begreppet extrakromosomalt cirkulärt DNA (eccDNA) blir alltså allt vanligare, medan termen dubbelminuter nu är reserverad för en specifik subtyp av eccDNA.
Strukturera
eccDNA är cirkulärt DNA som har hittats i mänskliga, växt- och djurceller och som finns i cellkärnan förutom det kromosomala DNA :t . eccDNA kan särskiljas från annat cirkulärt DNA i celler, såsom mitokondriellt DNA (mtDNA), eftersom det varierar i storlek från några hundra baser till megabaser och härrör från genomiskt DNA. Till exempel kan eccDNA bildas från exoner av proteinkodande gener, som mucin och titin . Forskare har antagit att eccDNA kan bidra till uttrycket av olika isoformer av en gen genom att interferera med eller främja transkriptionen av specifika exoner .
eccDNA har klassificerats som en av fyra olika kategorier av cirkulärt DNA baserat på storlek och sekvens, inklusive små polydispergerat cirkulärt DNA (spcDNA), telomera cirklar (t-cirklar), mikroDNA (100-400 bp) och extrakromosomalt DNA ( ecDNA ) . Var och en av dessa typer har sina egna unika biologiska egenskaper (se tabell 2):
| Namn på eccDNA | Storlek | Egenskaper | Fungera |
|---|---|---|---|
| spcDNA | 100–10 kb | Mycket olika typer av eccDNA, det finns ett stort intervall av antalet spcDNA-celler som hittats | Inblandad i mänsklig genetisk instabilitet |
| Telomeriska cirklar | multiplar av 738 bp | Bildas av telomera arrayer, som är en serie upprepade sekvenser i slutet av linjärt DNA. | Inblandad i alternativ förlängning av telomerer (ALT) |
| mikroDNA | 100-400 bp | från genomiska platser som har ett högt GC-innehåll och exondensitet | Uttryck små funktionella regulatoriska RNA (t.ex. mikroRNA och nya ärliknande RNA). |
| ecDNA | 1-3 Mb | Inkludera fullständiga gener, inga telomerer , acentriska | Amplifiera gener involverade i utvecklingen av cancer och läkemedelsresistens |
eccDNA-biogenes
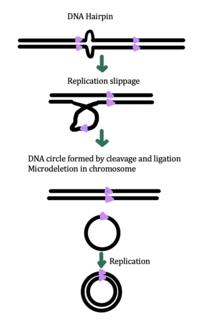


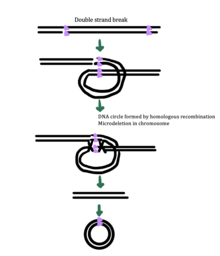
Även om den exakta mekanismen för eccDNA-generering fortfarande är okänd, har vissa studier föreslagit att eccDNA-generering kan vara kopplad till reparation av DNA-skador, hypertranskription, homolog rekombination och replikationsstress . Det finns flera föreslagna mekanismer för eccDNA-bildning: (1) replikationsglidning skapar en loop på mallsträngen som sedan skärs ut och ligeras in i en cirkel och lämnar en mikrodeletion på kromosomen , (2) replikationsglidning skapar en loop i produktsträngen som skärs ut och ligeras in i en cirkel som inte genererar en mikrodeletion i kromosomen , (3) ODERA-mekanismen för eccDNA-bildning, och (4) ett dubbelsträngat brott i en upprepad region repareras genom homolog rekombination , under vilken fragmentet bildas en cirkel och kromosomen drabbas av en mikrodeletion
Forskning utförd 2021 visade att apoptotiska celler är en källa till eccDNA; detta drogs slutsatsen på grund av studien som visar att apoptotisk DNA-fragmentering (ADF) är en förutsättning för eccDNA-bildning genom reningsmetoder.
eccDNA i icke-cancerceller
För att testa om eccDNA förekommer i icke-cancerceller, användes embryonala stamceller från mus och Southern Blot- analys; resultaten bekräftade att eccDNA finns i både cancerceller och icke-cancerceller. Det är också känt att eccDNA sannolikt inte kommer från specifika genomregioner; sekvenseringsdata från 2021 rapporterar att data tyder på att eccDNA är utbredda över hela genomet . Genomkartläggning av eccDNAs i full längd visade deras olika genomiska anpassningsmönster, vilket inkluderar vid intilliggande, överlappande eller kapslade positioner på samma kromosom eller över olika kromosomer . eccDNA härstammar mestadels från enstaka, kontinuerliga genomiska loci, vilket betyder att ett enda genomiskt fragment självcirkulerar för att bilda eccDNA:t, snarare än att bildas från ligering av olika genomiska fragment. Dessa två varianter kan klassificeras som kontinuerliga respektive icke-kontinuerliga eccDNAs. För att ytterligare förstå orsaken bakom cirkulariseringen av fragmenterat DNA, testades de Lig1 , Lig3 tre olika däggdjursligasenzymerna : och Lig4 . Genom att använda knockout-modeller i CH12F3-mus B-lymfocytcellinjen , identifierade forskning som utfördes 2021 Lig3 som det huvudsakliga ligaset för eccDNA-generering i dessa celler.
Fungera
Den exakta funktionen av eccDNA har diskuterats, men vissa studier har föreslagit att eccDNA kan bidra till genamplifiering vid cancer , immunfunktion och åldrande .
eccDNA-funktion i immunsystemet
Enligt forskning utförd 2021 är en annan funktion av eccDNA:er deras roll som möjliga immunstimulerande medel . eccDNA inducerar signifikant typ I-interferoner (IFNa, IFNβ), interleukin-6 (IL-6) och tumörnekrosfaktor (TNF), ännu mer än linjärt DNA och andra generellt potenta cytokininducerare vid sina högsta koncentrationsnivåer. Liknande mönster observeras med makrofager eftersom data visade att eccDNA är mycket potenta immunstimulerande medel för att aktivera både benmärgshärledda dendritiska celler och benmärgshärledda makrofager . Dessutom förändrade experiment eccDNA-strukturen med ett nick per eccDNA-segment och behandlades därefter med enzymer för att generera linjära versioner av eccDNA. I dessa experiment cytokintranskription , en viktig markör för immunsystemets aktivitet, vara mycket högre i det obehandlade eccDNA jämfört med den linjäriserade behandlingen, vilket ger att den cirkulära strukturen hos eccDNA snarare än den genetiska sekvensen i sig ger eccDNA dess immun funktion.
eccDNA-funktion vid cancer
Några kända funktioner av eccDNA inkluderar bidrag till intercellulär genetisk heterogenitet i tumörer , och mer specifikt amplifieringen av onkogener och läkemedelsresistenta gener . Detta stöder också att generna på eccDNA uttrycks. Sammantaget har eccDNA kopplats till cancer och läkemedelsresistens , åldrande , genkompensation, och av denna anledning fortsätter det att vara ett viktigt diskussionsämne.
Ansökningar
Roll i cancer
En subtyp av eccDNA, såsom ecDNA, ribosomalt DNA- lokus ( extrakromosomalt rDNA-cirkel ) och dubbla minuter har associerats med genomisk instabilitet . Dubbelminuters ecDNA är fragment av extrakromosomalt DNA , som ursprungligen observerades i ett stort antal mänskliga tumörer inklusive bröst, lungor, äggstockar, tjocktarm och framför allt neuroblastom . De är en manifestation av genamplifiering under utvecklingen av tumörer, vilket ger cellerna selektiva fördelar för tillväxt och överlevnad. Dubbla minuter , som faktiska kromosomer , består av kromatin och replikerar i cellens kärna under celldelning . Till skillnad från typiska kromosomer är de sammansatta av cirkulära fragment av DNA , upp till bara några miljoner baspar i storlek och innehåller inga centromerer eller telomerer .
Dubbla minutkromosomer (DM), som presenteras som parade kromatinkroppar under ljusmikroskopi , har visat sig vara en undergrupp av ecDNA. Kromosomer med dubbla minuter representerar cirka 30 % av det cancerinnehållande spektrumet av ecDNA, inklusive enstaka kroppar, och har visat sig innehålla identiskt geninnehåll som enstaka kroppar. ecDNA-notationen omfattar alla former av det stora geninnehållande extrakromosomala DNA som finns i cancerceller . Denna typ av ecDNA ses vanligtvis i cancerceller med olika histologier , men praktiskt taget aldrig i normal vävnad. ecDNA tros produceras genom dubbelsträngsbrott i kromosomer eller överreplikation av DNA i en organism.
Den cirkulära formen av ecDNA skiljer sig från den linjära strukturen hos kromosomalt DNA på meningsfulla sätt som påverkar cancerpatogenesen . Onkogener som kodas på ecDNA har massiv transkriptionell produktion, rankad i den översta 1% av generna i hela transkriptomet . I motsats till bakteriella plasmider eller mitokondrie-DNA , kromatiniseras ecDNA och innehåller höga nivåer av aktiva histonmärken , men en brist på repressiva histonmärken. ecDNA- kromatinarkitekturen saknar den högre ordningens kompaktering som finns på kromosomalt DNA och är bland det mest tillgängliga DNA:t i hela cancergenomet.
Från eccDNA visade sig matrisfästeregioner (MAR) aktivera amplifiering av onkogener . Transfektion av dessa MARs in i humana embryonala njure 293T-celler resulterade i en ökning av genuttryck , vilket tyder på att dessa eccDNA-härledda MARs är involverade i onkogenaktivering. eccDNA verkar också spela en roll i andra cancerformer såsom bröstcancer , där onkogener i human epidermal tillväxtfaktorreceptor 2 (HER2)-positiva bröstcancergener i eccDNA amplifieras. Detta eccDNA har också visat förmågan att förvärva resistens mot terapier för receptortyrosinkinaser ( RTK), som HER26.
Roll i åldrandet
Jäst är modellorganismer för att studera åldrande , och eccDNA har visat sig ackumuleras i gamla celler och spelar en roll för att orsaka åldrande i jäst. Spekulationerna fortsätter om allmänheten av detta koncept i högre arter, som däggdjur .
Se även
Vidare läsning
- Sinclair DA, Guarente L (december 1997). "Extrakromosomala rDNA-cirklar - en orsak till åldrande i jäst" . Cell . 91 (7): 1033–1042. doi : 10.1016/s0092-8674(00)80493-6 . PMID 9428525 .
- Cohen S, Houben A, Segal D (mars 2008). "Extrakromosomalt cirkulärt DNA härlett från tandem upprepade genomiska sekvenser i växter". Planttidningen . 53 (6): 1027–1034. doi : 10.1111/j.1365-313x.2007.03394.x . PMID 18088310 .
- Cohen S, Regev A, Lavi S (februari 1997). "Liten polydispergerad cirkulär DNA (spcDNA) i mänskliga celler: association med genomisk instabilitet" . Onkogen . 14 (8): 977–985. doi : 10.1038/sj.onc.1200917 . PMID 9050997 .
- Kumar P, Dillon LW, Shibata Y, Jazaeri AA, Jones DR, Dutta A (september 2017). "Normala och cancerösa vävnader frisätter extrakromosomalt cirkulärt DNA (eccDNA) i cirkulationen" . Molekylär cancerforskning . 15 (9): 1197–1205. doi : 10.1158/1541-7786.MCR-17-0095 . PMC 5581709 . PMID 28550083 .
- Baskin F, Rosenberg RN, Dev V (juni 1981). "Korrelation av dubbelminuters kromosomer med instabil multidrug korsresistens i upptagsmutanter av neuroblastomceller" . Proceedings of the National Academy of Sciences of the United States of America . 78 (6): 3654–3658. Bibcode : 1981PNAS...78.3654B . doi : 10.1073/pnas.78.6.3654 . PMC 319629 . PMID 6943568 . Gratis fulltext.
- Barker PE (februari 1982). "Dubbla minuter i mänskliga tumörceller". Cancergenetik och cytogenetik . 5 (1): 81–94. doi : 10.1016/0165-4608(82)90043-7 . PMID 6175392 .
- Masters J, Keeley B, Gay H, Attardi G (maj 1982). "Variabelt innehåll av dubbelminuters kromosomer är inte korrelerat med graden av fenotypinstabilitet i metotrexatresistenta mänskliga cellinjer" . Molekylär och cellulär biologi . 2 (5): 498–507. doi : 10.1128/MCB.2.5.498 . PMC 369819 . PMID 7110138 . Gratis fulltext.