Angomonas deanei
|
|
| Angomonas deanei | |
|---|---|
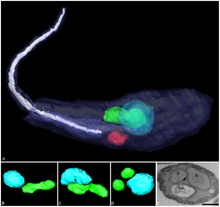
| Tredimensionell rekonstruktion av Angomonas deanei innehållande en bakteriell endosymbiont (grön) nära dess kärna (blå). | |
| Vetenskaplig klassificering | |
| Provins: | Euglenozoa |
| Klass: | Kinetoplastea |
| Beställa: | Trypanosomatida |
| Släkte: | Angomonas |
| Arter: |
A. deanei
|
| Binomialt namn | |
|
Angomonas deanei (Carvalho, 1973) Teixeira & Camargo, 2011 |
|
| Synonymer | |
|
Crithidia deanei Carvalho, 1973 |
|
Angomonas deanei är en flagellerad trypanosomatidprotozo . Som en obligatorisk parasit infekterar den mag-tarmkanalen hos insekter och är i sin tur en värd för symbiotiska bakterier. Den bakteriella endosymbiont Ca. " Kinetoplastibacterium crithidii " upprätthåller en permanent mutualistisk relation med protozoen så att den inte längre kan fortplanta sig och överleva på egen hand. Symbiosen, som senare också upptäckts i varierande grad hos andra protister som Strigomonas culicis , Novymonas esmeraldas , Diplonema japonicum och Diplonema aggregatum anses vara bra modeller för förståelsen av eukaryoternas utveckling från prokaryoter och cellorganellers ursprung (dvs. symbiogenes ).
Arten beskrevs första gången som Crithidia deanei 1973 av en brasiliansk parasitolog Aurora LM Carvalho. En fylogenetisk analys 2011 visade att den tillhör släktet Angomonas och blev därigenom Angomonas deanei . Den symbiotiska bakterien är en medlem av β-proteobakterien som härstammar från den gemensamma förfadern med släktet Bordetella , eller mer troligt, Taylorella . De två organismerna har varit beroende av varandra så mycket att bakterien inte kan föröka sig och protozoerna kan inte längre infektera insekter när de är isolerade.
Upptäckt
Angomonas deanei beskrevs ursprungligen som Crithidia deanei . 1973 upptäckte en brasiliansk doktorand Aurora Luiza de Moura Carvalho vid Universidade Federal de Goiás arten från sin studie av tarmparasiter från lönnmördaren i Goiás. Nästa år rapporterade han att insekten Zelus leucogrammus som han upptäckte inte var naturligt infekterad av protozoen, utan den förvärvades från andra insekter. Samtidigt rapporterade ett forskarlag vid Universidade de Brasilia de biokemiska egenskaperna och strukturella detaljerna baserat på transmissionselektronmikroskopi. De upptäckte att den hyser en endosymbiont och beskrev den som "förmodligen bakteriell" som tillhandahöll de "trypanosomatida essentiella näringsämnena." Endosymbiontens bakteriella natur bekräftades 1977 när det visades att den kunde dödas genom att behandlas med ett antibiotikum kloramfenikol och att det hjälper värden att syntetisera aminosyran arginin från ornitin.
När fler strukturella och molekylära detaljer studerades, blev skillnaden mellan A. deanei och andra Crithidia -arter mer uttalad. 1991 föreslog Maria Auxiliadora de Sousa och Suzana Corte-Real vid Instituto Oswaldo Cruz ett nytt släkte Angomonas för arten. Filogenetisk studie av Marta MG Teixeira och Erney P. Camargo vid universitetet i São Paulo med deras medarbetare 2011 validerade det nya artnamnet A. deanei tillsammans med en beskrivning av en ny besläktad art A. ambiguus , som också innehåller samma bakteriella endosymbiont .
Strukturera

Kroppen av Angomonas deanei är elliptisk till formen, med en framträdande svansliknande flagell vid dess bakre ände för rörelse. Den bakteriella endosymbionten finns inuti sin kropp och är omgiven av två cellmembran som är typiska för gramnegativa bakterier , men dess cellmembran uppvisar ovanliga egenskaper, såsom närvaron av fosfatidylkolin , en huvudmembranlipid (atypisk för bakteriemembran) och den mycket reducerat peptidoglykanskikt , som visar minskad eller frånvaro av stel cellvägg . Cellmembranet hos protozovärden innehåller en 18-domän β-fat porin , som är ett karakteristiskt protein för gramnegativa bakterier och ovanligt för eukaryoter. Dessutom innehåller den kardiolipin och fosfatidylkolin som de viktigaste fosfolipiderna , medan steroler är frånvarande. Kardiolipin är en typisk lipid av bakteriella membran; fosfatidylkolin, å andra sidan, är mest närvarande i symbiotiska prokaryoter av eukaryota celler. För symbiotisk anpassning har den protozoiska värden genomgått förändringar såsom reducerad paraflagellär stav, vilket krävs för full motilitet hos bakterieflagellerna. Ändå är den paraflagella stavgenen PFR1 fullt fungerande. Den saknar också introner och transkription av långa polycistroniska mRNA som krävs av andra eukaryoter för komplexa genaktiviteter. Hela genomet är fördelat i 29 kromosomer och innehåller 10 365 proteinkodande gener, 59 överförings-RNA, 26 ribosomala RNA och 62 icke-kodande RNA.
Medan protozoen har sina separata mitokondrier som tillhandahåller elektrontransportsystem för produktion av cellulär energi, produceras ATP- molekylerna genom dess glykosomer . Bakterien är känd för att tillhandahålla viktiga näringsämnen till värden. Den syntetiserar aminosyror, vitaminer, kvävehaltiga baser och hem för protozoerna. Hem är nödvändigt för protozoernas tillväxt och utveckling. Bakterien tillhandahåller också enzymerna för ureacykeln som saknas i värden. I gengäld erbjuder protozoen sina enzymer för de fullständiga metaboliska vägarna för biosyntesen av aminosyror , lipider och nukleotider , som saknas i bakterien. Bakterien har kraftigt reducerat genom jämfört med dess besläktade bakteriearter, och saknar många gener som är nödvändiga för dess överlevnad. Fosfatidylinositol , en membranlipid som krävs för cell-cell-interaktion i bakterierna, syntetiseras också av protozoen. Bakterien är också beroende av värden för ATP-molekyler för dess energiska funktioner. Således delar de två organismerna intimt och utbyter sina metaboliska system.
När bakterien dödas med antibiotika kan protozoen inte längre infektera insekter, på grund av den förändrade glykosylfosfatidylinositol (gp63) i protozoens flagellum. En bakteriefri protozo uppvisar minskade genaktiviteter; särskilt de som är involverade i oxidationsreduktionsprocessen, ATP-hydrolyskopplad protontransport och glykolys stoppas. De strukturella komponenterna förändras också inklusive cellyta, kolhydratsammansättning, paraflagellar stav och kinetoplast.
Parasitism
Angomonas deanei upptäcktes ursprungligen från matsmältningskanalen hos insekten Zelus leucogrammus . Men man insåg att insekterna inte är kraftigt infekterade och troligen överfördes från andra insekter. Det är nu känt att infektera olika myggor och flugor och kan infektera fibroblastceller från däggdjur under experimentella förhållanden. Överföring från en insekt till en annan sker endast mellan vuxna ( horisontell överföring ), och protozoen kan inte fixera sig i baktarmen på insektslarver. Flagellen används som ett adhesivt organ som fästs nära rektalkörtlarna och någon gång direkt på ytan av rektalkörtlarna.
Fortplantning
Den cellulära reproduktionen visar en stark synergistisk anpassning mellan bakterien och protozoan. Bakterien delar sig först, följt av protozoernas organeller och sist kärnan . Som ett resultat innehåller dotterprotozoerna exakt samma kopior av organellerna och den bakteriella endosymbionten. Hela reproduktionen tar cirka 6 timmar i ett idealiskt odlingsmedium; Således kan en enda protozo producera 256 dotterceller på en dag, även om den kan skilja sig något under sin naturliga livsmiljö.
Endosymbiont och evolution
Symbiotiska bakterier i trypanosomatida protozoer härstammar från en β-proteobakterie. Med A. deanei , bakterierna Ca. " Kinetoplastibacterium crithidii " har utvecklats tillsammans i ett mutualistiskt förhållande som kännetecknas av intensiva metaboliska utbyten. Endosymbiont innehåller enzymer och metaboliska prekursorer som fullbordar väsentliga biosyntetiska vägar för värdprotozoen, såsom de i ureacykeln och produktionen av hemin och polyamin .
Den symbiotiska bakterien tillhör β-proteobacterium-familjen Alcaligenaceae . Baserat på 16S rRNA- gensekvenserna är det känt att det härstammar från en gemensam förfader med den i Strigomonas culicis . De två grupperna antas gå in i två olika värdprotozoer för att utvecklas till olika arter. gavs det vetenskapliga namnet ( Candidatus ) Kinetoplastibacterium crithidii till bakterien. Även om det ursprungligen föreslogs att bakterien utvecklats från en gemensam förfader med medlemmar av Bordetella , visade detaljerad fylogenomisk analys att den är närmare besläktad med medlemmar av släktet Taylorella . Omanalys av GTDB finner släktets syster till Proftella , en symbiont av Diaphorina citri .