Dominans (genetik)

Inom genetik är dominans fenomenet av en variant ( allel ) av en gen på en kromosom som maskerar eller åsidosätter effekten av en annan variant av samma gen på den andra kopian av kromosomen . Den första varianten kallas dominant och den andra recessiv . Detta tillstånd att ha två olika varianter av samma gen på varje kromosom orsakas ursprungligen av en mutation i en av generna, antingen ny ( de novo ) eller ärvd . Termerna autosomal dominant eller autosomal recessiv används för att beskriva genvarianter på icke-könade kromosomer ( autosomer ) och deras associerade egenskaper, medan de på könskromosomer (allosomer) benämns X-linked dominant, X -linked recessive eller Y-linked ; dessa har ett arv och presentationsmönster som beror på både förälderns och barnets kön (se Könskoppling ) . Eftersom det bara finns en kopia av Y-kromosomen kan Y-kopplade egenskaper inte vara dominanta eller recessiva. Dessutom finns det andra former av dominans som ofullständig dominans , där en genvariant har en partiell effekt jämfört med när den finns på båda kromosomerna, och samdominans , där olika varianter på varje kromosom båda visar sina associerade egenskaper.
Dominans är ett nyckelbegrepp inom mendelskt arv och klassisk genetik . Bokstäver och Punnett-rutor används för att demonstrera principerna för dominans i undervisningen, och användningen av stora bokstäver för dominanta alleler och små bokstäver för recessiva alleler är en allmänt följd konvention. Ett klassiskt exempel på dominans är nedärvningen av fröform i ärter . Ärter kan vara runda, associerade med allel R eller rynkiga, associerade med allel r . I detta fall är tre kombinationer av alleler (genotyper) möjliga: RR , Rr och rr . RR ( homozygota )-individerna har runda ärtor, och rr -individerna (homozygota) har rynkiga ärtor . Hos Rr ( heterozygota ) individer maskerar R -allelen närvaron av r -allelen, så dessa individer har också runda ärter. Således är allelen R dominant över allelen r , och allelen r är recessiv till allelen R.
Dominans är inte inneboende i en allel eller dess egenskaper ( fenotyp) . Det är en strikt relativ effekt mellan två alleler av en given gen av vilken funktion som helst; en allel kan vara dominant över en andra allel av samma gen, recessiv till en tredje och samdominant med en fjärde. Dessutom kan en allel vara dominant för en egenskap men inte för andra.
Dominans skiljer sig från epistasis , fenomenet med en allel av en gen som maskerar effekten av alleler av en annan gen.
Bakgrund
_(14582377398).jpg)
Begreppet dominans introducerades av Gregor Johann Mendel . Även om Mendel, "genetikens fader", först använde termen på 1860-talet, var den inte allmänt känd förrän i början av nittonhundratalet. Mendel observerade att för en mängd olika egenskaper hos trädgårdsärter som har att göra med utseendet på frön, fröskidor och växter, fanns det två diskreta fenotyper, såsom runda kontra skrynkliga frön, gula mot gröna frön, röda mot vita blommor eller höga kontra korta växter. När de odlades separat producerade växterna alltid samma fenotyper, generation efter generation. Men när linjer med olika fenotyper korsades (interbred), dök en och endast en av föräldrafenotyperna upp hos avkomman (grön, eller rund, eller röd eller lång). Men när dessa hybridväxter korsades visade avkomplantorna de två ursprungliga fenotyperna, i ett karakteristiskt 3:1-förhållande, varvid den vanligaste fenotypen var föräldrahybridväxterna. Mendel resonerade att varje förälder i den första korsningen var en homozygot för olika alleler (en förälder AA och den andra föräldern aa), att var och en bidrog med en allel till avkomman, med resultatet att alla dessa hybrider var heterozygoter (Aa), och att en av de två allelerna i hybridkorset dominerade uttrycket av den andra: A maskerad a. Den sista korsningen mellan två heterozygoter (Aa X Aa) skulle producera AA-, Aa- och aa-avkommor i ett genotypförhållande på 1:2:1 där de två första klasserna visar (A)-fenotypen och den sista visar (a)-fenotypen 3:1 fenotypförhållandet.
Mendel använde inte termerna gen, allel, fenotyp, genotyp, homozygot och heterozygot, som alla introducerades senare. Han introducerade notationen av stora och små bokstäver för dominanta respektive recessiva alleler, som fortfarande används idag.
1928 föreslog den brittiske befolkningsgenetikern Ronald Fisher att dominansen agerade baserat på naturligt urval genom bidraget från modifierande gener . År 1929 svarade den amerikanske genetikern Sewall Wright med att konstatera att dominans helt enkelt är en fysiologisk konsekvens av metabola vägar och den relativa nödvändigheten av den involverade genen. Wrights förklaring blev ett faktum inom genetiken, och debatten avslutades i stort sett. Vissa egenskaper kan dock ha sin dominans påverkad av evolutionära mekanismer.
Kromosomer, gener och alleler
De flesta djur och vissa växter har parade kromosomer och beskrivs som diploida . De har två versioner av varje kromosom, en från moderns ägg och den andra från faderns sperma , känd som könsceller , beskriven som haploid och skapad genom meios . Dessa könsceller smälter sedan samman under befruktning under sexuell reproduktion , till en ny encellig zygot , som delar sig flera gånger, vilket resulterar i en ny organism med samma antal kromosompar i varje (icke-gamet) cell som dess föräldrar. Inom däggdjursgenetik har autosomala dominanta störningar stamtavlor som visar ett vertikalt mönster av arv.
Varje kromosom i ett matchande (homologt) par är strukturellt likt det andra och har en mycket liknande DNA- sekvens ( loci , singular locus). DNA:t i varje kromosom fungerar som en serie diskreta gener som påverkar olika egenskaper. Således har varje gen också en motsvarande homolog, som kan existera i olika versioner som kallas alleler . Allelerna på samma lokus på de två homologa kromosomerna kan vara identiska eller olika.
Till exempel bestäms blodtypen hos människor av ABO-genen som kodar för varianter av ett enzym som skapar blodgruppen A, B, AB eller O som finns på den långa eller q-armen av kromosom nio (9q34.2). Det finns tre olika alleler som kan vara närvarande på detta ställe, men bara två kan vara närvarande i vilken individ som helst, en ärvd från sin mor och en från sin far.
Om två alleler av en given gen är identiska, kallas organismen en homozygot och sägs vara homozygot med avseende på den genen; om istället de två allelerna är olika är organismen en heterozygot och är heterozygot. Den genetiska sammansättningen av en organism, antingen på ett enda lokus eller över alla dess gener tillsammans, kallas dess genotyp . Genotypen av en organism, direkt och indirekt, påverkar dess molekylära, fysiska och andra egenskaper, som individuellt eller kollektivt kallas dess fenotyp . Vid heterozygota genlägen interagerar de två allelerna för att producera fenotypen. [ citat behövs ]
Typer av dominans
Fullständig dominans
Vid fullständig dominans maskerar effekten av en allel i en heterozygot genotyp helt effekten av den andra. Allelen som maskerar anses vara dominant för den andra allelen, och den maskerade allelen anses vara recessiv .
Fullständig dominans i en heterozygots fenotyp går inte att skilja från en dominant homozygots fenotyp. Ett klassiskt exempel på fullständig dominans är nedärvningen av fröform (ärtform) i ärter. Ärter kan vara runda (associerade med allel R ) eller skrynkliga (associerade med allel r ). I detta fall är tre kombinationer av alleler ( genotyper ) möjliga: RR, rr, Rr. RR och rr är homozygota och Rr är heterozygota. RR - individerna har runda ärtor och rr -individerna har skrynkliga ärtor. Hos Rr- individer maskerar R -allelen närvaron av r -allelen, så dessa individer har också runda ärtor. Således är allelen R fullständigt dominant för allelen r och allelen r är recessiv till allelen R.
Ofullständig dominans

Ofullständig dominans (även kallad partiell dominans , semi-dominans , intermediär nedärvning , eller ibland felaktigt samdominans i reptilgenetik) uppstår när fenotypen av den heterozygota genotypen är skild från och ofta mellanliggande till fenotyperna av den homozygota genotypen. Det fenotypiska resultatet uppträder ofta som en blandad form av egenskaper i heterozygot tillstånd. Till exempel snapdragonblommans färg homozygot för antingen röd eller vit. När den röda homozygota blomman paras med den vita homozygota blomman, ger resultatet en rosa snapdragon-blomma. Den rosa snapdragon är resultatet av ofullständig dominans. En liknande typ av ofullständig dominans finns i växten klockan fyra, där rosa färg produceras när äkta uppfödda föräldrar till vita och röda blommor korsas. Inom kvantitativ genetik , där fenotyper mäts och behandlas numeriskt, om en heterozygots fenotyp ligger exakt mellan (numeriskt) den av de två homozygoterna, sägs fenotypen inte uppvisa någon dominans alls, dvs dominans existerar endast när heterozygotens fenotyp mäter fenotypens fenotyp. till en homozygot än den andra.
När växter av F 1 -generationen självpollineras kommer det fenotypiska och genotypiska förhållandet för F 2 -generationen att vara 1:2:1 (Röd: Rosa: Vit).
Se hypotes om partiell dominans .
Meddominans



Samdominans uppstår när bidragen från båda allelerna är synliga i fenotypen och ingen av allelen maskerar en annan.
IA Till exempel, i ABO-blodgruppssystemet kontrolleras kemiska modifieringar av ett glykoprotein (H-antigenet) på ytan av blodkroppar av tre alleler, varav två är co-dominanta till varandra ( , I B ) och dominant över det recessiva i vid ABO-lokuset . I A- allelerna och I B- producerar olika modifieringar. Enzymet som I A kodar för lägger till en N-acetylgalaktosamin till ett membranbundet H-antigen. I B - enzymet tillför en galaktos. i - allelen producerar ingen modifiering. Således I A- och I B -allelerna var och en dominant för i ( I A I A och I A i -individer har båda typ A-blod och I B I B- och I B i- individer har båda typ B-blod), men I A I B- individer har båda modifieringar på sina blodkroppar och har således blod av typ AB, så I A- och I B -allelerna sägs vara samdominanta.
Ett annat exempel förekommer vid platsen för beta-globinkomponenten i hemoglobin , där de tre molekylära fenotyperna Hb A /Hb A , Hb A /Hb S och Hb S /Hb S alla kan särskiljas genom proteinelektrofores . (Det medicinska tillståndet som produceras av den heterozygota genotypen kallas sickle-cell trait och är ett lindrigare tillstånd som kan skiljas från sickle-cell anemi , således visar allelerna ofullständig dominans med avseende på anemi, se ovan). För de flesta genloci på molekylär nivå uttrycks båda allelerna co-dominant, eftersom båda transkriberas till RNA .
Samdominans, där allelprodukter samexisterar i fenotypen, skiljer sig från ofullständig dominans, där den kvantitativa interaktionen av allelprodukter producerar en mellanliggande fenotyp. Till exempel, i samdominans kommer en röd homozygot blomma och en vit homozygot blomma att producera avkomma som har röda och vita fläckar. När växter av F1-generationen är självpollinerade, kommer det fenotypiska och genotypiska förhållandet för F2-generationen att vara 1:2:1 (Röd:Fläckig:Vit). Dessa förhållanden är desamma som för ofullständig dominans. Återigen är denna klassiska terminologi olämplig – i verkligheten bör sådana fall inte alls sägas uppvisa dominans.
Att ta itu med vanliga missuppfattningar
Dominans relaterar till förhållandet mellan två versioner av en gen. En dominerande egenskap är vanligtvis i överensstämmelse med arvsmönster som kan ses i Punnett Squares. Om en individ har två versioner av en gen, anses genen som ofta observeras i ytterligare generationer vara "dominant".
Inom genetiken finns det några missuppfattningar som är ganska vanliga. Man tror att en dominant egenskap är "starkare" och "övermannar" en recessiv egenskap. Dominanta egenskaper antas också vara mer benägna att ärvda såväl som vanligare i en population. Idén om att dominerande egenskaper är manliga eller maskulina är en annan vanlig missuppfattning. Uppkomsten av dessa olika idéer beror på de olika begreppen dominans i icke-genetiska miljöer; som att vara stark, kraftfull och kontrollerande; som skiljer sig från det genetiska begreppet dominans.
Dominans avgör inte om en allel är skadlig, neutral eller fördelaktig. Selektion måste dock verka på gener indirekt genom fenotyper och dominans påverkar exponeringen av alleler i fenotyper, därav förändringshastigheten i allelfrekvenser under selektion. Skadliga recessiva alleler kan kvarstå i en population vid låga frekvenser, med de flesta kopior som bärs i heterozygoter, utan kostnad för dessa individer. Dessa sällsynta recessiva är grunden för många ärftliga genetiska sjukdomar .
Nomenklatur
Inom genetiken började symboler som algebraiska platshållare. När en allel är dominant för en annan är den äldsta konventionen att symbolisera den dominanta allelen med en stor bokstav. Den recessiva allelen tilldelas samma bokstav med små bokstäver. I ärtexemplet, när dominansförhållandet mellan de två allelerna är känt, är det möjligt att beteckna den dominanta allelen som ger en rund form med en stor bokstavssymbol R och den recessiva allelen som ger en skrynklig form med en lägre- fallsymbol r . De homozygota dominanta, heterozygota och homozygota recessiva genotyperna skrivs sedan RR , Rr respektive rr . Det skulle också vara möjligt att beteckna de två allelerna som W och w , och de tre genotyperna WW , Ww och ww , av vilka de två första producerade runda ärtor och den tredje rynkiga ärtor. Valet av " R " eller " W " som symbol för den dominanta allelen bedömer inte i förväg om allelen som orsakar den "runda" eller "rynkiga" fenotypen när homozygot är den dominanta.
En gen kan ha flera alleler. Varje allel symboliseras av lokussymbolen följt av en unik upphöjd. Hos många arter är den vanligaste allelen i den vilda populationen betecknad vildtypsallelen. Det symboliseras med ett +-tecken som en upphöjd. Andra alleler är dominanta eller recessiva till vildtypsallelen. För recessiva alleler är lokussymbolen med små bokstäver. För alleler med någon grad av dominans till vildtypsallelen är den första bokstaven i lokussymbolen med versaler. Till exempel, här är några av allelerna vid laboratoriemusens a- lokus, Mus musculus : A y , dominant gul; a + , vildtyp; och en bt , black and tan. a bt - allelen är recessiv mot vildtypsallelen och Ay . -allelen är samdominant till vildtypsallelen Ay- allelen . är också samdominant till a bt -allelen, men visar att förhållandet ligger utanför gränserna för reglerna för genetisk nomenklatur för mus
Regler för genetisk nomenklatur har utvecklats i takt med att genetiken har blivit mer komplex. Kommittéer har standardiserat reglerna för vissa arter, men inte för alla. Reglerna för en art kan skilja sig något från reglerna för en annan art.
Förhållande till andra genetiska begrepp
Flera alleler
Även om varje individ av en diploid organism har högst två olika alleler på vilket lokus som helst (förutom aneuploidier ), finns de flesta gener i ett stort antal allelversioner i populationen som helhet. Om allelerna har olika effekter på fenotypen kan ibland deras dominansförhållanden beskrivas som en serie.
Till exempel påverkas pälsfärgen hos tamkatter av en serie alleler av TYR -genen (som kodar för enzymet tyrosinas ). Allelerna C , cb av pigment och följaktligen olika nivåer , cs och ca av (fullfärgade, burmesiska , siamesiska respektive albino ) producerar olika nivåer färgutspädning . C - allelen (fullfärg) är helt dominant över de tre sista och c a -allelen (albino) är helt recessiv mot de tre första.
Autosomal kontra könsbunden dominans
Hos människor och andra däggdjursarter bestäms kön av två könskromosomer som kallas X-kromosomen och Y - kromosomen . Mänskliga honor är XX ; män är XY . De återstående kromosomparen finns hos båda könen och kallas autosomer ; genetiska egenskaper associerade med loci på dessa kromosomer beskrivs som autosomala och kan vara dominanta eller recessiva. Genetiska egenskaper på X- och Y -kromosomerna kallas könsbundna, eftersom de är kopplade till könskromosomer, inte för att de är karakteristiska för det ena eller det andra könet. I praktiken hänvisar termen nästan alltid till X -länkade egenskaper och väldigt många sådana egenskaper (som röd-grön färgseendebrist) påverkas inte av sex. Honor har två kopior av varje genlokus som finns på X-kromosomen, precis som för autosomerna, och samma dominansförhållanden gäller. Hanar har dock bara en kopia av varje X-kromosomgenlokus och beskrivs som hemizygota för dessa gener. Y-kromosomen är mycket mindre än X och innehåller en mycket mindre uppsättning gener, inklusive, men inte begränsat till, de som påverkar "manlighet", såsom SRY- genen för testikelbestämmande faktor . Dominansregler för könsbundna genloci bestäms av deras beteende hos honan: eftersom hanen bara har en allel (förutom i fallet med vissa typer av Y-kromosomaneuploidi ), uttrycks den allelen alltid oavsett om den är dominant eller recessiv. Fåglar har kromosomer av motsatt kön: hanfåglar har ZZ och honfåglar ZW kromosomer. Men nedärvning av egenskaper påminner XY-systemet på annat sätt; zebrafinkar hanar kan bära vit färgande gen i sin ena av två Z-kromosomer, men honor utvecklar alltid vit färg. Gräshoppor har XO-system. Kvinnor har XX, men män bara X. Det finns ingen Y-kromosom alls.
Epistasis
Epistasis [" epi + stasis = att sitta på toppen"] är en interaktion mellan alleler vid två olika genloci som påverkar en enskild egenskap, som ibland kan likna en dominansinteraktion mellan två olika alleler på samma locus. Epistasis modifierar det karakteristiska 9:3:3:1 som förväntas för två icke-epistatiska gener. För två loci erkänns 14 klasser av epistatiska interaktioner. Som ett exempel på recessiv epistas kan ett genlokus avgöra om ett blompigment är gult ( AA eller Aa ) eller grönt ( aa ), medan ett annat lokus avgör om pigmentet produceras ( BB eller Bb ) eller inte ( bb ). I en bb -växt kommer blommorna att vara vita, oavsett genotypen av det andra lokuset som AA , Aa eller aa . bb - kombinationen är inte dominant för A- allelen: snarare visar B -genen recessiv epistas till A -genen, eftersom B - lokuset när det är homozygott för den recessiva allelen ( bb ) undertrycker fenotypiskt uttryck av A -lokuset. I en korsning mellan två AaBb -växter ger detta ett karakteristiskt förhållande på 9:3:4 , i detta fall av gula : gröna : vita blommor.
I dominant epistas kan ett genlokus bestämma gult eller grönt pigment som i föregående exempel: AA och Aa är gula och aa är gröna. Ett andra lokus bestämmer om en pigmentprekursor produceras ( dd ) eller inte ( DD eller Dd ). Här, i en DD- eller Dd -växt, kommer blommorna att vara färglösa oberoende av genotypen på A -lokuset, på grund av den epistatiska effekten av den dominanta D -allelen. I en korsning mellan två AaDd -växter kommer alltså 3/4 av växterna att vara färglösa, och de gula och gröna fenotyperna uttrycks endast i dd -växter. Detta ger ett karakteristiskt 12:3:1 av vita : gula : gröna växter.
Kompletterande epistas uppstår när två loci påverkar samma fenotyp. Till exempel, om pigmentfärg produceras av CC eller Cc men inte cc , och av DD eller Dd men inte dd , produceras inte pigment i någon genotypisk kombination med antingen cc eller dd . Det vill säga, båda loci måste ha minst en dominant allel för att producera fenotypen. Detta ger ett karakteristiskt 9:7 mellan pigmenterade och opigmenterade växter. Komplementär epistas däremot producerar en opigmenterad växt om och endast om genotypen är cc och dd , och det karakteristiska förhållandet är 15:1 mellan pigmenterade och opigmenterade växter.
Klassisk genetik betraktade epistatiska interaktioner mellan två gener åt gången. Det är nu uppenbart från molekylär genetik att alla genloci är involverade i komplexa interaktioner med många andra gener (t.ex. metabola vägar kan involvera mängder av gener), och att detta skapar epistatiska interaktioner som är mycket mer komplexa än de klassiska två-lokusmodellerna .
Hardy–Weinberg-principen (uppskattning av bärvågsfrekvens)
Frekvensen av det heterozygota tillståndet (som är bärartillståndet för en recessiv egenskap) kan uppskattas med hjälp av Hardy–Weinbergs formel :

Denna formel gäller för en gen med exakt två alleler och relaterar frekvensen av dessa alleler i en stor population till frekvenserna för deras tre genotyper i den populationen.
Till exempel , om p är frekvensen för allelen A och q är frekvensen av allelen a så är termerna p2 , 2pq och q2 frekvenserna för genotyperna AA , Aa respektive aa . Eftersom genen bara har två alleler måste alla alleler vara antingen A eller a och p + q = 1 . Om A nu är helt dominant för a så kan inte frekvensen av bärargenotypen Aa observeras direkt (eftersom den har samma egenskaper som den homozygota genotypen AA ), men den kan uppskattas utifrån frekvensen av den recessiva egenskapen i populationen , eftersom detta är samma som för den homozygota genotypen aa . dvs de individuella allelfrekvenserna kan uppskattas: q = √ f (aa) , p = 1 − q , och från dessa kan frekvensen för bärargenotypen härledas: f (Aa) = 2 pq .
Denna formel bygger på ett antal antaganden och en korrekt uppskattning av frekvensen av det recessiva draget. I allmänhet kommer alla verkliga situationer att avvika från dessa antaganden till viss del, vilket kommer att införa motsvarande felaktigheter i uppskattningen. Om den recessiva egenskapen är sällsynt, kommer det att vara svårt att uppskatta dess frekvens exakt, eftersom en mycket stor provstorlek kommer att behövas.
Dominant kontra fördelaktig
Egenskapen "dominant" förväxlas ibland med begreppet fördelaktig och egenskapen "recessiv" förväxlas ibland med begreppet skadlig, men fenomenen är distinkta. Dominans beskriver fenotypen av heterozygoter med avseende på homozygoternas fenotyper och utan hänsyn till i vilken grad olika fenotyper kan vara fördelaktiga eller skadliga. Eftersom många genetiska sjukdomsalleler är recessiva och eftersom ordet dominans har en positiv klang, görs ofta antagandet att den dominerande fenotypen är överlägsen med avseende på kondition. Detta är dock inte garanterat; som diskuteras nedan medan de flesta genetiska sjukdomsalleler är skadliga och recessiva, är inte alla genetiska sjukdomar recessiva.
Ändå har denna förvirring varit genomgående genom genetikens historia och kvarstår till denna dag. Att ta itu med denna förvirring var en av de främsta motiven för publiceringen av Hardy–Weinberg-principen .
Molekylära mekanismer
Den molekylära grunden för dominans var okänd för Mendel. Det är nu underförstått att ett genlokus inkluderar en lång serie (hundratals till tusentals) av baser eller nukleotider av deoxiribonukleinsyra (DNA) vid en speciell punkt på en kromosom. Den centrala dogmen inom molekylärbiologi säger att " DNA gör RNA gör protein ", det vill säga att DNA transkriberas för att göra en RNA-kopia, och RNA översätts för att göra ett protein. I denna process kan olika alleler på ett lokus transkriberas eller inte, och om de transkriberas kan de översättas till något olika versioner av samma protein (kallade isoformer ). Proteiner fungerar ofta som enzymer som katalyserar kemiska reaktioner i cellen, som direkt eller indirekt producerar fenotyper. Mutationer inom genomet kan förändra den katalytiska aktiviteten och därför påverka dominansen. I vilken diploid organism som helst kan DNA-sekvenserna för de två alleler som finns närvarande vid vilket genställe som helst vara identiska (homozygota) eller olika (heterozygota). Även om genlokuset är heterozygot på nivån av DNA-sekvensen, kan proteinerna som görs av varje allel vara identiska. I avsaknad av någon skillnad mellan proteinprodukterna kan ingen av allelen sägas vara dominant (se samdominans ovan). Även om de två proteinprodukterna är något olika ( allozymer ), är det troligt att de producerar samma fenotyp med avseende på enzymverkan, och återigen ingen av allelerna kan sägas vara dominanta.
Zygositet
antog Mendels lag om oberoende sortiment att alleler kommer att sorteras oberoende, med en allel som "dominant". Zygositet , graden av likhet mellan en organisms alleler, kan påverka dominansen. Inom en diploid organism skulle dessa definieras av allelernas haplotypinteraktioner . Genhaploidi kan resultera i att en enda funktionell allel gör tillräckligt med protein för att producera en fenotyp som är identisk med den hos homozygoten . Tre allmänna typer av haplotypinteraktioner är möjliga:
- Haplosufficiens. I en diploid skulle en funktionell allel av en haplosufficient gen anses vara dominant, medan en icke-funktionell allel skulle anses vara recessiv. Anta till exempel att standardmängden enzym som produceras i den funktionella homozygoten är 100 %, med de två funktionella allelerna som bidrar med 50 % vardera. Den enda funktionella allelen i heterozygoten producerar 50 % av standardmängden enzym, vilket är tillräckligt för att producera standardfenotypen. Om heterozygoten och den funktionella allelhomozygoten har identiska fenotyper, är den funktionella allelen dominant för den icke-funktionella allelen. Detta sker vid albinogenen: heterozygoten producerar tillräckligt med enzym för att omvandla pigmentprekursorn till melanin, och individen har standardpigmentering. Till exempel, hos människor och andra organismer, uppstår den opigmenterade huden av albinofenotypen när en individ är homozygot för en allel som kodar för en icke-funktionell version av ett enzym som behövs för att producera hudpigmentet melanin .
- Ofullständig Haploinsufficiens. Mindre vanligt ger närvaron av en enda funktionell allel en fenotyp som inte är normal, men mindre allvarlig, än den för den icke-funktionella homozygoten. Detta inträffar när den funktionella allelen inte är haplotillräcklig, så termerna haploinsufficiens och ofullständig dominans används vanligtvis i dessa fall. Den mellanliggande interaktionen inträffar där den heterozygota genotypen producerar en fenotyp mellan de två homozygoterna. Beroende på vilken av de två homozygoterna heterozygoten liknar mest, sägs den ena allelen visa ofullständig dominans över den andra. Till exempel, hos människor Hb- genlokuset ansvarigt för Beta-kedjans protein ( HBB ) som är ett av de två globinproteiner som utgör blodpigmentet hemoglobin . Många människor är homozygota för en allel som kallas Hb A ; vissa personer bär på en alternativ allel som kallas HbS , antingen som homozygoter eller heterozygoter. Hemoglobinmolekylerna av Hb S / Hb S -homozygoter genomgår en formförändring som förvränger de röda blodkropparnas morfologi och orsakar en allvarlig, livshotande form av anemi som kallas sicklecellanemi . Personer som är heterozygota Hb A / Hb S för denna allel har en mycket mindre allvarlig form av anemi som kallas sickle-cell-egenskap . Eftersom sjukdomsfenotypen av Hb A / Hb S heterozygoter är mer lik men inte identisk med Hb A / Hb A -homozygoten, sägs Hb A -allelen vara ofullständigt dominant för Hb S -allelen.
- Fullständig haploinsufficiens. En enda funktionell allel i heterozygoten kan producera otillräcklig genprodukt för någon funktion av genen, vilket gör att de vanligtvis icke-funktionella allelerna blir dominerande. Fenotypen kommer då att likna den hos en homozygot med icke-funktionell allel istället för vildtypen. Den icke-funktionella allelen skulle sägas vara dominant för vildtypsfenotypens funktionella allel. Denna situation kan uppstå när den icke-funktionella allelen producerar ett defekt protein som stör den korrekta funktionen av proteinet som produceras av standardallelen. Närvaron av det defekta proteinet "dominerar" standardproteinet, och sjukdomsfenotypen hos heterozygoten liknar mer den hos homozygoten för två defekta alleler. Termen "dominant" används ofta felaktigt för defekta alleler vars homozygota fenotyp inte har undersökts, men som orsakar en distinkt fenotyp när de är heterozygota med den normala allelen. Detta fenomen förekommer i ett antal upprepade trinukleotidsjukdomar , ett exempel är Huntingtons sjukdom . Vid Huntingtons sjukdom orsakar fullständig haploinsufficiens den dominerande effekten av mutantproteinet. Normalt har en person cirka 20 CAG-nukleotidupprepningar i sin HTT-gen, men de med Huntingtons sjukdom har 40+ CAG-repetitioner. Ett annat exempel är Marfans syndrom, en ärftlig bindvävssjukdom, orsakad av en mutation i fibrillin-1 (FBN1) genen. En normal kopia av FBN1-genen ärvs från en förälder medan en dominant onormal FBN1-genkopia ärvs av en annan förälder.
Dominant-negativa mutationer
Många proteiner är normalt aktiva i form av en multimer, ett aggregat av flera kopior av samma protein, annars känt som ett homomultimert protein eller homoligomert protein . Faktum är att en majoritet av de 83 000 olika enzymerna från 9800 olika organismer i BRENDA Enzyme Database representerar homoligomerer. När vildtypsversionen av proteinet är närvarande tillsammans med en mutantversion kan en blandad multimer bildas. En mutation som leder till ett mutant protein som stör aktiviteten av vildtypsproteinet i multimeren är en dominant-negativ mutation.
En dominant-negativ mutation kan uppstå i en human somatisk cell och ge en proliferativ fördel för mutantcellen, vilket leder till dess klonala expansion. Till exempel kan en dominant-negativ mutation i en gen som är nödvändig för den normala processen av programmerad celldöd ( apoptos ) som svar på DNA-skada göra cellen resistent mot apoptos. Detta kommer att tillåta proliferation av klonen även när överdriven DNA-skada är närvarande. Sådana dominant-negativa mutationer förekommer i tumörsuppressorgenen p53 . P53 vildtypsproteinet är normalt närvarande som en fyra-protein multimer (oligotetramer). Dominant-negativa p53- mutationer förekommer i ett antal olika typer av cancer och pre-cancerösa lesioner (t.ex. hjärntumörer, bröstcancer, orala pre-cancerösa lesioner och oral cancer).
Dominant-negativa mutationer förekommer även i andra tumörsuppressorgener. Till exempel identifierades två dominant-negativa könslinjemutationer i Ataxia telangiectasia mutated ( ATM) som ökar känsligheten för bröstcancer. Dominanta negativa mutationer av transkriptionsfaktorn C/EBPα kan orsaka akut myeloid leukemi. Ärftliga dominanta negativa mutationer kan också öka risken för andra sjukdomar än cancer. Dominant-negativa mutationer i Peroxisome proliferator-activated receptor gamma ( PPARγ) är associerade med allvarlig insulinresistens, diabetes mellitus och hypertoni.
Dominant-negativa mutationer har också beskrivits i andra organismer än människor. Faktum är att den första studien som rapporterade ett mutantprotein som hämmade den normala funktionen av ett vildtypsprotein i en blandad multimer var med bakteriofag T4 svansfiberproteinet GP37. Mutationer som producerar ett trunkerat protein snarare än ett mutant protein av full längd verkar ha den starkaste dominant-negativa effekten i studierna av P53, ATM, C/EBPα och bakteriofag T4 GP37.

Ärftliga defekter i enzymer ärvs i allmänhet på ett autosomalt sätt eftersom det finns fler icke-X-kromosomer än X-kromosomer, och ett recessivt sätt eftersom enzymerna från de opåverkade generna i allmänhet är tillräckliga för att förhindra symtom hos bärare. Undantag inkluderar fall av haploinsufficiens , där den opåverkade genen inte kan kompensera för den drabbade.
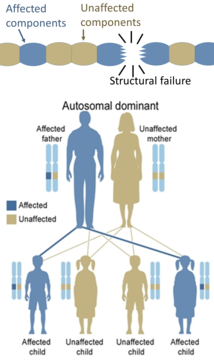
Å andra sidan är ärftliga defekter i strukturella proteiner (såsom osteogenesis imperfecta , Marfans syndrom och Ehlers–Danlos syndrom ) generellt autosomalt dominanta, eftersom det räcker med att vissa komponenter är defekta för att hela strukturen ska bli dysfunktionell. Detta är en dominant-negativ process, där en muterad genprodukt negativt påverkar den icke-muterade genprodukten inom samma cell.


Dominanta och recessiva genetiska sjukdomar hos människor
Hos människor klassificeras många genetiska egenskaper eller sjukdomar helt enkelt som "dominanta" eller "recessiva". Speciellt med så kallade recessiva sjukdomar, som visserligen är en faktor för recessiva gener, men som kan förenkla den underliggande molekylära grunden och leda till missförstånd av dominansens natur.
Till exempel är den recessiva genetiska sjukdomen fenylketonuri (PKU) ett resultat av någon av ett stort antal (>60) alleler vid genlokuset för enzymet fenylalaninhydroxylas ( PAH ). Många av dessa alleler producerar lite eller ingen PAH , som ett resultat av vilket substratet fenylalanin (Phe) och dess metaboliska biprodukter ackumuleras i det centrala nervsystemet och kan orsaka allvarliga intellektuella funktionshinder om de inte behandlas.
För att illustrera dessa nyanser visas genotyperna och de fenotypiska konsekvenserna av interaktioner mellan tre hypotetiska PAH-alleler i följande tabell:
| Genotyp | PAH -aktivitet | [ Phe ] konc | PKU ? |
|---|---|---|---|
| AA | 100 % | 60 μM | Nej |
| AB | 30 % | 120 μM | Nej |
| CC | 5 % | 200 ~ 300 μM | Hyperfenylalaninemi |
| BB | 0,3 % | 600 ~ 2400 μM | Ja |
Hos opåverkade personer som är homozygota för en standard funktionell allel ( AA ), är PAH- aktivitet standard (100%), och koncentrationen av fenylalanin i blodet [ Phe ] är cirka 60 μM (= μmol/L ). Hos obehandlade personer som är homozygota för en av PKU-allelerna ( BB ), är PAH- aktiviteten nära noll, [Phe] tio till fyrtio gånger standard, och individen manifesterar PKU.
I AB- heterozygoten är PAH- aktiviteten endast 30 % (inte 50 %) av standarden, blod [ Phe ] är dubbelt förhöjd och personen uppvisar inte PKU. Således A- allelen dominant för B -allelen med avseende på PKU, men B -allelen är ofullständigt dominant för A -allelen med avseende på dess molekylära effekt, bestämning av PAH- aktivitetsnivå (0,3 % < 30 % << 100 %) . Slutligen A -allelen ofullständigt dominant för B -allelen med avseende på [Phe], eftersom 60 μM < 120 μM << 600 μM. Observera ännu en gång att det är irrelevant för frågan om dominans att den recessiva allelen producerar en mer extrem [Phe] fenotyp.
För en tredje allel C producerar en CC- homozygot en mycket liten mängd PAH- enzym, vilket resulterar i en något förhöjd nivå av [ Phe ] i blodet, ett tillstånd som kallas hyperfenylalaninemi , vilket inte resulterar i intellektuell funktionsnedsättning.
Det vill säga dominansförhållandena för två alleler kan variera beroende på vilken aspekt av fenotypen som övervägs. Det är vanligtvis mer användbart att prata om de fenotypiska konsekvenserna av de allelinteraktioner som är involverade i någon genotyp, snarare än att försöka tvinga in dem i dominanta och recessiva kategorier.
Se även
- Ambidirectional dominans
- Lista över Mendelska egenskaper hos människor
- Mitokondriellt DNA
- Punnett square
- "On-line anteckningar för Biology 2250 – Principles of Genetics" . Memorial University of Newfoundland.
- Online Mendelian Inheritance in Man (OMIM): Hemoglobin—Beta Locus; HBB - 141900 — Sickle-cellanemi
- Online Mendelian Inheritance in Man (OMIM): ABO Glycosyltransferase - 110300 — ABO blodgrupper