Anatomi av lillhjärnan
| Lillhjärnan | |
|---|---|
 Ritning av den mänskliga hjärnan, visar lillhjärnan och pons
| |

Vertikal mittlinjetvärsnitt av den mänskliga lillhjärnan, visar vikningsmönster för cortex och inre strukturer
| |
| Detaljer | |
| Del av | Metencephalon |
| Artär | SCA , AICA , PICA |
| Ven | överlägsen , underlägsen |
| Identifierare | |
| NeuroLex ID | birnlex_1489 |
| TA98 | A14.1.07.001 |
| TA2 | 5788 |
| Anatomiska termer för neuroanatomi | |
Lillhjärnans anatomi kan ses på tre nivåer. På nivån av grov anatomi består lillhjärnan av ett hårt vikt och skrynkligt lager av cortex, med vit substans under, flera djupa kärnor inbäddade i den vita substansen och en vätskefylld kammare i mitten . På mellannivå kan lillhjärnan och dess hjälpstrukturer delas upp i flera hundra eller tusen självständigt fungerande moduler eller fack som kallas mikrozoner . På mikroskopisk nivå består varje modul av samma lilla uppsättning neuronala element, upplagda med en mycket stereotyp geometri.
Grov anatomi
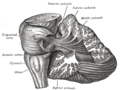
Lillhjärnan ligger vid basen av hjärnan, med den stora massan av hjärnbarken ovanför den och den del av hjärnstammen som kallas pons framför den. Den är separerad från den överliggande storhjärnan av ett lager av seg dura mater ; alla dess förbindelser med andra delar av hjärnan färdas genom pons. Anatomer klassificerar lillhjärnan som en del av metencephalon , som också inkluderar pons; metencephalon i sin tur är den övre delen av rhombencephalon eller "bakhjärnan". Liksom hjärnbarken är lillhjärnan uppdelad i två hemisfärer; den innehåller också en smal mittlinjezon som kallas vermis . En uppsättning stora veck används konventionellt för att dela upp den övergripande strukturen i tio mindre lobuler .
På grund av dess stora antal små granulceller innehåller lillhjärnan fler neuroner än resten av hjärnan tillsammans, men den tar bara upp 10% av den totala hjärnvolymen. Lillhjärnan tar emot nästan 200 miljoner ingående fibrer; däremot består synnerven av bara en miljon fibrer.
Det ovanliga utseendet på lillhjärnan döljer det faktum att huvuddelen av strukturen består av ett mycket tätt vikt lager av grå substans, lillhjärnbarken . Det har uppskattats att om den mänskliga hjärnbarken kunde vecklas ut helt skulle den ge upphov till ett lager av neural vävnad som är cirka 1 meter lång och 10 centimeter bred – en total yta på 500-1000 kvadratcm, allt packat inom en volym av 100-150 kubik cm. Under den grå substansen i cortex ligger vit substans , som till stor del består av myeliniserade nervfibrer som löper till och från cortex. Inbäddade i den vita substansen - som ibland kallas arbor vitae (Livets träd) i lillhjärnan på grund av dess grenade, trädliknande utseende - finns fyra djupa cerebellära kärnor .
Lillhjärnan kan delas in enligt tre olika kriterier: grovanatomisk, fylogenetisk och funktionell.
Grova anatomiska uppdelningar
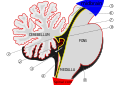
Vid grov inspektion kan tre lober urskiljas i lillhjärnan: den flockulonodulära loben , den främre loben (rostralt till "primärfissuren") och den bakre loben (dorsal till "primärfissuren"). De två sistnämnda kan ytterligare delas in i en mittlinje cerebellar vermis och laterala cerebellära hemisfärer.

Figur 3: Cerebellum och omgivande regioner; sagittal vy av en halvklot. S: Mellanhjärnan . B: Pons . C: Medulla . D: Ryggmärg . E: Fjärde ventrikeln . F: Arbor vitae . G: Tonsil . H: Främre lob. I: Bakre lob.
|

Fylogenetiska och funktionella indelningar
Lillhjärnan kan också delas in i tre delar baserat på både fylogenetiska kriterier (den evolutionära åldern för varje del) och på funktionella kriterier (de inkommande och utgående kopplingar varje del har och den roll som spelas för normal cerebellär funktion). Från den fylogenetiskt äldsta till den nyaste är de tre delarna:
| Funktionell valör ( fylogenetisk valör) | Anatomiska delar | Roll |
| Vestibulocerebellum (Archicerebellum) | Flockulonodulär lob (och omedelbart intilliggande vermis) | Vestibulocerebellum reglerar balans och ögonrörelser. Den tar emot vestibulär input från både de halvcirkulära kanalerna och från de vestibulära kärnorna och skickar fibrer tillbaka till de mediala och laterala vestibulära kärnorna. Den får också visuell input från de överlägsna colliculi och från den visuella cortex (den senare via pontinkärnorna , som bildar en cortico-ponto-cerebellär bana). Lesioner i vestibulocerebellum orsakar balans- och gångstörningar . Det finns en annan liten region, känd som biventer lobule . |
| Spinocerebellum (Paleocerebellum) | Vermis och mellanliggande delar av hemisfärerna ("paravermis") | Spinocerebellum reglerar kroppens och extremiteternas rörelser. Den tar emot proprioceptioninput från ryggmärgens ryggpelare (inklusive spinocerebellarkanalen ) och trigeminusnerven , såväl som från syn- och hörselsystem . Det skickar fibrer till djupa cerebellära kärnor (inklusive den fastigiala kärnan ) som i sin tur skjuter ut till både hjärnbarken (via mellanhjärnan och thalamus ) och hjärnstammen (via retikulär bildning i pons och vestibulära kärnor i medulla oblongata ), alltså tillhandahåller modulering av fallande motorsystem. Spinocerebellum innehåller sensoriska kartor när det tar emot data om positionen för olika kroppsdelar i rymden: i synnerhet tar vermis fibrer från bålen och proximala delar av extremiteterna, medan de mellanliggande delarna av hemisfärerna tar emot fibrer från de distala delarna av extremiteterna. . Spinocerebellum kan utveckla proprioceptiv input för att förutse den framtida positionen för en kroppsdel under loppet av en rörelse, på ett "feed forward" sätt. |
| Cerebrocerebellum (Neocerebellum, Pontocerebellum) | Laterala delar av hemisfärerna | Neohjärnan är involverad i att planera rörelse och utvärdera sensorisk information för handling. Den tar endast emot input från hjärnbarken (särskilt parietalloben ) via pontinkärnorna (i pons , bildar cortico-ponto-cerebellära vägar) och dentate nucleus (i lillhjärnan ), och skickar fibrer huvudsakligen till den ventrolaterala thalamus (i) sväng kopplad till motoriska områden i den premotoriska cortex och primära motoriska delen av hjärnbarken) och till den röda kärnan (i sin tur kopplad till den nedre olivarkärnan , som länkar tillbaka till de cerebellära hemisfärerna). Neohjärnan är involverad i planering av rörelser som är på väg att inträffa och har också rent kognitiva funktioner. |
Mycket av vad man förstår om lillhjärnans funktioner härrör från noggrann dokumentation av effekterna av fokala lesioner hos mänskliga patienter som har lidit av skada eller sjukdom eller genom forskning om djurskador.
Cellulär anatomi
Som förklaras mer i detalj i avsnittet Funktion , skiljer sig lillhjärnan från de flesta andra hjärnområden genom att flödet av neurala signaler genom det är nästan helt enkelriktat: det finns praktiskt taget inga bakåtkopplingar mellan dess neuronala element. Således är det mest logiska sättet att beskriva den cellulära strukturen att börja med ingångarna och följa sekvensen av anslutningar till utgångarna.
Djupa kärnor
De fyra djupa kärnorna i lillhjärnan är de dentata , emboliforma , klotformade och fastigii kärnorna och de fungerar som de huvudsakliga kommunikationscentrumen, sänder och tar emot information till och från specifika delar av hjärnan. Dessutom tar dessa kärnor emot både hämmande och excitatoriska signaler från andra delar av hjärnan som i sin tur påverkar kärnans utgående signaler.(De klotformiga och emboliformiga kärnorna utgör den mellanliggande kärnan ).
Kortikala lager


Cytoarkitekturen ( cellulär organisation) av lillhjärnan är mycket enhetlig, med anslutningar organiserade i en grov, tredimensionell uppsättning av vinkelräta kretselement . Denna organisatoriska enhetlighet gör att nervkretsarna är relativt lätta att studera.
Det finns tre lager till cerebellar cortex; från yttre till inre skikt, dessa är de molekylära, Purkinje- och granulära skikten. Cerebellarbarkens funktion är i huvudsak att modulera information som flödar genom de djupa kärnorna. Lillhjärnans mikrokretsar är schematiserade i figur 5. Mossiga och klättrande fibrer bär sensorimotorisk information in i de djupa kärnorna, som i sin tur skickar den vidare till olika premotoriska områden, vilket reglerar förstärkningen och timingen av motoriska handlingar. Mossiga och klättrande fibrer matar också in denna information i hjärnbarken, som utför olika beräkningar, vilket resulterar i reglering av Purkinje-cellsbränning. Purkinje-neuroner matas tillbaka in i de djupa kärnorna via en potent hämmande synaps . Denna synaps reglerar i vilken utsträckning mossiga och klättrande fibrer aktiverar de djupa kärnorna, och kontrollerar därmed lillhjärnans yttersta effekt på motorfunktionen. Den synaptiska styrkan hos nästan varje synaps i cerebellar cortex har visat sig genomgå synaptisk plasticitet . Detta gör det möjligt för kretsarna i cerebellarbarken att kontinuerligt justera och finjustera utsignalen från cerebellum, vilket utgör grunden för vissa typer av motorisk inlärning och koordination. Varje lager i cerebellarbarken innehåller de olika celltyperna som utgör denna krets.
Molekylärt lager
Detta yttersta skikt av lillhjärnbarken innehåller två typer av hämmande interneuroner : stjärnceller och korgceller . Den innehåller också Purkinje-neuronernas dendritiska arbor och parallella fiberkanaler från granulcellerna. Både stellat och korgceller bildar GABAergiska synapser på Purkinje-cellsdendriter.
Purkinje lager
Det mellersta lagret innehåller bara en typ av cellkropp – den av den stora Purkinje-cellen . Purkinjeceller är de primära integrerande neuronerna i cerebellarbarken och ger dess enda uteffekt. Purkinjecellsdendriter är stora berså med hundratals taggiga grenar som når upp i det molekylära lagret (fig. 6). Dessa dendritiska arbors är platta - nästan alla av dem ligger i plan - med närliggande Purkinje arbors i parallella plan. Varje parallell fiber från granulcellerna löper ortogonalt genom dessa arbors, som en tråd som passerar genom många lager. Purkinjeneuroner är GABAergiska – vilket betyder att de har hämmande synapser – med nervcellerna i de djupa cerebellära och vestibulära kärnorna i hjärnstammen. Varje Purkinje-cell får excitatorisk input från 100 000 till 200 000 parallella fibrer. Parallella fibrer sägs vara ansvariga för den enkla (allt eller inget, amplitudinvariant ) spikning av Purkinje-cellen.
Purkinjeceller får också input från den nedre olivärkärnan via klättringsfibrer . Ett bra minnesmärke för denna interaktion är frasen "klättra upp i det andra olivträdet", med tanke på att klättringsfibrer härstammar från den kontralaterala sämre olivoljan. I slående kontrast till de över 100 000 ingångarna från parallella fibrer, får varje Purkinje-cell input från exakt en klättringsfiber; men denna enda fiber "klättrar" upp i Purkinjecellens dendriter, slingrar sig runt dem och gör ett stort antal synapser allt eftersom. Nettoinmatningen är så stark att en enda aktionspotential från en klätterfiber kan producera en "komplex spik" i Purkinje-cellen: en skur av flera spikar i rad, med minskande amplitud, följt av en paus under vilken enkla spikar är undertryckta.
Strax under Purkinje-skiktet finns Lugaro-cellerna vars mycket långa dendriter färdas längs gränsen mellan Purkinje- och de granulära skikten.
Granulärt lager
Det innersta lagret innehåller cellkropparna av tre typer av celler: de många och små granulcellerna , de något större unipolära borstcellerna och de mycket större Golgi-cellerna . Mossiga fibrer kommer in i det granulära lagret från deras huvudsakliga ursprung, pontinkärnorna. Dessa fibrer bildar excitatoriska synapser med granulcellerna och cellerna i de djupa cerebellära kärnorna. Granulcellerna skickar sina T-formade axoner – kända som parallella fibrer – upp i det ytliga molekylära lagret, där de bildar hundratusentals synapser med Purkinje- celldendriter . Den mänskliga lillhjärnan innehåller i storleksordningen 60 till 80 miljarder granulceller, vilket gör denna encellstyp till den överlägset mest talrika neuronen i hjärnan (ungefär 70 % av alla neuroner i hjärnan och ryggmärgen, tillsammans). Golgi-celler ger hämmande återkoppling till granulceller, bildar en synaps med dem och projicerar ett axon in i det molekylära lagret.
Relation med hjärnbarken
De lokala fältpotentialerna hos neocortex och lillhjärnan svänger koherent vid (6–40 Hz) hos vakna djur. Dessa verkar vara under kontroll av produktionen från hjärnbarken. Denna utmatning skulle förmedlas av en väg från lager 5/6 neuroner i neocortex genom det projektet antingen till pons eller den sämre oliven. Om genom pons detta skulle gå till mossiga fibrer som synapsar med granulat och Golgi neuroner med granulcellerna sedan rikta Purkinje neuroner via deras excitatoriska parallella fibrer. Om den sämre olivoljan skulle gå via excitatoriska klättringsfiberingångar till Purkinje-neuroner. Dessa returnerar denna utmatning tillbaka till hjärnbarken genom den ventrolaterala thalamus som avslutar slingan.
Den kortikopontocerebellära vägen är den största vägen förknippad med lillhjärnan. Dessa fibrer, som uppstår i hjärnbarken, slutar först ipsilateralt i pontinkärnorna . Sedan dekusserar fibrerna och bildar den mellersta cerebellarskaftet, som slutar i cerebellar cortex som mossiga fibrer. Denna väg sänder signaler som informerar lillhjärnan om den pågående rörelsen och den kommande rörelsen. Detta hjälper den kontinuerliga justeringen av motorisk aktivitet.
Initieringen av rörelsen vidarebefordras till lillhjärnan via corticoreticulocerebellar-banan. Dessa synapsar ipsilateralt i den retikulära formationen , sedan via de nedre och mellersta peduncles in i cerebellar vermis .
Den motoriska cortex och somatosensoriska cortex projicerar ut på de ipsilaterala nedre och accessoriska olivärkärnorna och bildar sedan den olivocerebellära kanalen . Cortico-olivarfibrer synapsar bilateralt i den nedre olivarkärnan . Ordningen bevaras i de olivocerebellära traktens projektioner på "kroppskartorna" i den kontralaterala cerebellar cortex. Under vilande förhållanden i djurförsök, urladdas grupper av olivära neuroner synkront vid 5 till 10 Hz (impulser/s). I cerebellarbarken tar Purkinje-cellernas respons i form av komplexa spikar.
Lillhjärnan skickar sina projektioner tillbaka till hjärnbarken via lillhjärnan .
Den cerebellära laterala expansionen, eller neocerebellum, kan vara associerad med kognitiva funktioner, och den är anatomiskt kopplad till den laterala prefrontala cortex . Den visar störst aktivitet under tal, med en ensidig övervikt som överensstämmer med en möjlig koppling (via thalamus) med det motoriska talområdet.
När lesioner uppstår i de associationsområden som är kopplade till lillhjärnan av kortikopontocerebellära fibrer, kan det kognitiva affektiva syndromet uppstå. Detta resulterar i kognitiva defekter i form av minskad resonemangskraft, ouppmärksamhet, grammatiska fel i tal, dålig rumslig känsla och ojämn minnesförlust.
Blodtillförsel


Tre artärer förser lillhjärnan med blod (fig. 7): den övre cerebellära artären (SCA), den främre inferior cerebellarartären (AICA) och den bakre cerebellära artären (PICA).
SCA förgrenar sig från den laterala delen av basilarartären, precis under dess bifurkation in i den bakre cerebrala artären . Här lindar den sig baktill runt pons (till vilken den också förser blod) innan den når lillhjärnan. SCA levererar blod till det mesta av cerebellar cortex, cerebellar nuclei och de superior cerebellar peduncles.
AICA förgrenar sig från den laterala delen av basilarartären, precis ovanför korsningen mellan de vertebrala artärerna. Från sitt ursprung förgrenar den sig längs den nedre delen av pons vid lillhjärnans vinkel innan den når lillhjärnan. Denna artär tillför blod till den främre delen av den nedre lillhjärnan, den mellersta cerebellarskaftet och till ansikts- ( CN VII) och vestibulokochleära nerver (CN VIII). Obstruktion av AICA kan orsaka pareser , förlamning och förlust av känsel i ansiktet; det kan också orsaka hörselnedsättning . Dessutom kan det orsaka en infarkt i cerebellopontinvinkeln. Detta kan leda till hyperakusi (dysfunktion av stapediusmuskeln , innerverad av CN VII ) och yrsel (fel tolkning från den vestibulära halvcirkulära kanalens endolymfaceleration orsakad av förändring av CN VIII ).
PICA förgrenar sig från den laterala delen av de vertebrala artärerna precis under deras förbindelse med basilarartären. Innan den når den nedre ytan av lillhjärnan skickar PICA grenar in i märgen och förser flera kranialnervkärnor med blod . I lillhjärnan förser PICA med blod till den bakre nedre delen av lillhjärnan, den nedre delen av lillhjärnan, nucleus ambiguus , den vagusmotoriska kärnan, den spinala trigeminuskärnan , den solitära kärnan och de vestibulocochleära kärnorna.
Variationer bland ryggradsdjur
Det finns stor variation i storleken och formen på lillhjärnan hos olika ryggradsdjursarter. Det är allmänt störst i brosk- och benfiskar , fåglar och däggdjur, men något mindre i reptiler. De stora parade och invecklade loberna som finns hos människor är typiska för däggdjur, men lillhjärnan är i allmänhet en enda medianlob i andra grupper och är antingen slät eller endast lätt räfflad. Hos däggdjur är neocerebellum huvuddelen av lillhjärnan i massa, men hos andra ryggradsdjur är det vanligtvis spinocerebellum.
Hos amfibier , lampreys och hagfish är lillhjärnan föga utvecklad; i de två sistnämnda grupperna är den knappt urskiljbar från hjärnstammen. Även om spinocerebellum finns i dessa grupper, är de primära strukturerna små parade kärnor som motsvarar vestibulocerebellum.
Peduncles
Lillhjärnan följer det allmänna mönstret för grupper av tre som finns i anatomin, med tre stora in- och utgående cerebellära pedunkler (fiberbuntar). Dessa är de överlägsna (brachium conjunctivum), mellersta (brachium pontis) och underlägsna (restiforma och juxtarestiforma kroppar ) cerebellära skaft.
| Stjälk | Beskrivning |
| Överlägsen | Även om det finns några afferenta fibrer från den främre spinocerebellarkanalen som transporteras till den främre lillhjärnloben via denna pedunkel, är de flesta fibrerna efferenter. Sålunda är den överlägsna cerebellära pelaren den huvudsakliga utgångsvägen för cerebellum. De flesta av de efferenta fibrerna har sitt ursprung i dentatkärnan som i sin tur skjuter ut till olika mellanhjärnans strukturer inklusive den röda kärnan , den ventrala laterala/ventrala främre kärnan i thalamus och medulla . Dentatorubrothalamocortical (dentate nucleus > röd kärna > thalamus > premotor cortex ) och cerebellothalamocortical ( cerebellum > thalamus > premotor cortex) är två huvudvägar som passerar genom denna peduncle och är viktiga i motorisk planering . |
| Mitten | Denna består helt och hållet av afferenta fibrer som har sitt ursprung i pontinkärnorna som en del av den massiva corticopontocerebellar-kanalen (hjärnbarken > pons > cerebellum). Dessa fibrer härstammar från de sensoriska och motoriska områdena i hjärnans neocortex och gör den mellersta cerebellära pelaren till den största av de tre cerebellära pelarna. |
| Sämre |
Detta bär många typer av in- och utgående fibrer som huvudsakligen sysslar med att integrera proprioceptiv sensorisk input med motoriska vestibulära funktioner såsom balans och hållning. Proprioceptiv information från kroppen transporteras till lillhjärnan via den dorsala spinocerebellarkanalen . Detta område passerar genom den nedre cerebellära pelaren och synapser i paleocerebellum. Vestibulär information projicerar på höljet. Klätterfibrerna i den inferior oliven löper genom den inferior cerebellar skaftet . Denna peduncle bär också information direkt från Purkinje-cellerna ut till de vestibulära kärnorna i den dorsala hjärnstammen som ligger vid korsningen mellan pons och medulla . |
Det finns tre källor för input till lillhjärnan, i två kategorier bestående av mossiga respektive klätterfibrer. Mossiga fibrer kan härröra från pontinkärnorna, som är kluster av neuroner som finns i pons som bär information från den kontralaterala hjärnbarken. De kan också uppstå i det spinocerebellära området vars ursprung ligger i den ipsilaterala ryggmärgen . Det mesta av utflödet från lillhjärnan synapserar initialt till de djupa cerebellära kärnorna innan det går ut via de tre pedunklerna. Det mest anmärkningsvärda undantaget är den direkta hämningen av de vestibulära kärnorna av Purkinje-celler.
Utveckling
Under de tidiga stadierna av embryonal utveckling , börjar hjärnan att bildas i tre distinkta segment: prosencephalon , mesencephalon och rhombencephalon . Rombencephalon är det mest kaudala (mot svansen) segmentet av den embryonala hjärnan; det är från detta segment som lillhjärnan utvecklas. Längs det embryonala rhombencefaliska segmentet utvecklas åtta svullnader, kallade rhombomerer . Lillhjärnan uppstår från två rhombomerer belägna i nervplattan i nervröret , en struktur som så småningom bildar hjärnan och ryggmärgen. De specifika rombomererna från vilka lillhjärnan bildas är rombomer 1 (Rh.1) kaudalt (nära svansen) och "isthmus" rostralt (nära fronten).
Två primära regioner tros ge upphov till nervcellerna som utgör lillhjärnan. Den första regionen är den ventrikulära zonen i taket på den fjärde ventrikeln . Detta område producerar Purkinje-celler och djupa cerebellära kärnneuroner . Dessa celler är de primära utgångsneuronerna i cerebellar cortex och cerebellum. Den andra könszonen (cellulär födelseplats) är känd som den rombiska läppen, neuroner flyttar sedan genom mänskliga embryonala vecka 27 till det yttre granulära lagret . Detta lager av celler - som finns på utsidan av lillhjärnan - producerar granulneuronerna. Granulneuronerna migrerar från detta yttre skikt för att bilda ett inre skikt känt som det inre granulskiktet. Det externa granulära lagret upphör att existera i den mogna lillhjärnan och lämnar endast granulceller kvar i det inre granullagret. Den vita delen av lillhjärnan kan vara en tredje groddzon i lillhjärnan; dess funktion som groddzon är dock kontroversiell.
Ytterligare bilder
Dissektion som visar projektionsfibrerna i lillhjärnan
Schema av tak av fjärde ventrikeln. Pilen är i Majendies foramen.

Mänsklig hjärna midsagittal vy
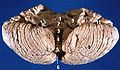
Framifrån av den mänskliga lillhjärnan, med siffror som indikerar framträdande landmärken




| Myndighetskontroll : Vetenskapliga databaser |
|---|