Waptia
|
Waptia Tidsintervall:
|
|
|---|---|

|
|

|
|
| Fossila exemplar av Waptia | |
| Artists rekonstruktion av Waptia fieldensis | |
| Vetenskaplig klassificering | |
| Rike: | Animalia |
| Provins: | Arthropoda |
| Clade : | Mandibulata |
| Beställa: | † Hymenocarina |
| Släkte: |
† Waptia Walcott, 1912 |
| Arter: |
† W. fieldensis
|
| Binomialt namn | |
|
† Waptia fieldensis
Walcott , 1912
|
|

|
|
| Placeringen av Burgess Shale -formationen i British Columbia | |
Waptia fieldensis är en utdöd artropod från Mellankambriska Burgess Shale Lagerstätte i Kanada . Den växte till en längd av 6,65 cm (3 tum) och hade ett stort skaldjur och en segmenterad kropp som slutade i ett par svansflikar. Det var en aktiv simmare och troligen ett rovdjur av mjukkroppar. Det är också ett av de äldsta djuren med direkta bevis på avelvård . Waptia fieldensis är den enda arten som klassificeras under släktet Waptia . Andra exemplar av Waptia är kända från Spence Shale i Utah.
Baserat på antalet individer är Waptia fieldensis den tredje vanligaste leddjuren från Burgess Shale Formation, med tusentals insamlade exemplar. Det var bland de första fossilerna som hittades av den amerikanske paleontologen Charles D. Walcott 1909. Han beskrev det 1912 och döpte det efter två berg nära upptäcktsplatsen – Wapta Mountain och Mount Field , andra exemplar
Även om den har en anmärkningsvärd likhet med moderna kräftdjur , var dess taxonomiska affiniteter länge oklara. En omfattande ombeskrivning publicerad 2018 klassificerade den som en medlem av Hymenocarina (som innehåller många andra tvåskaliga leddjur) inom Mandibulata .
Beskrivning

Kända exemplar av Waptia varierar i längd från 13,5 till 66,5 millimeter (0,53 till 2,62 tum) med de allra flesta (~85 %) som är 40 till 60 millimeter (1,6 till 2,4 tum) långa. Den tvåskaliga ryggskölden var sadelformad och var tunn och icke mineraliserad, och var troligen flexibel i livet. Ryggskölden komprimerades i sidled och hade ingen distinkt gräns mellan de två halvorna. Ryggskölden var endast fäst vid kroppen i en liten sektion nära framsidan av huvudet. Kroppen var uppdelad i tre huvudsegment, cephalothorax (huvud), post-cephalothorax och buken.
Framsidan av huvudet bar ett par reniforma (njurformade) ögon, cirka 1 millimeter (0,039 tum) i diameter, som föddes på korta stjälkar. Ett exemplar med bevarade ommatidia visar att densiteten av ommatidia i ögat var cirka 600 per kvadratmillimeter. Det föreslås att detta tillät god sido- och framåtseende. Ett par små lober, cirka 1 millimeter (0,039 tum) långa, sticker ut nära ögonen. Liknande strukturer är kända från de besläktade Canadaspis såväl som andra mandibulater, och tros motsvara kräftdjurens hemi-ellipsoida kroppar, och har därför sannolikt en luktfunktion . Mellan ögonen finns en triangulär struktur, kallad "median triangulär projektion", som förmodligen är homolog med den "anterior sclerite" hos andra kambriska leddjur. Huvudet bär ett par antenner, som är sammansatta av 10 långsträckta cylindriska segment/ podomerer , som sekventiellt minskar i bredd mot antennens spets. De främre ändarna av varje podomer bär setae , som är orienterade i en vinkel på 75° till 95° i förhållande till antennens axel.
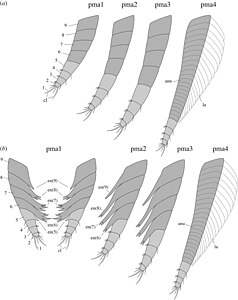
Mandibeln har ett tresegmenterat utsprång, som är täckta med setae . Underkäken visar tecken på sklerotisering mot kanten där de två underkäkarna kom i kontakt, som har en tandad marginal. Mandibeln hade sannolikt en bitande och malande funktion. Överkäken , ändpodomererna bär ett par klor, tillsammans med många setae. Dessa assisterade troligen matmanipulation vid sidan av underkäkarna. Cephalothorax har ytterligare fyra par uniramösa benliknande bihang, varav de tre första är väl segmenterade, med 5 endopodsegment , som är spetsade med klor, med en 4 eller 5 segmenterad basipod med välutvecklade änditer , särskilt på det första paret , som skjuter ut inåt från benen. Det fjärde benet skiljer sig i det faktum att endast den allra sista delen av benet är segmenterad, med resten annullerad, med de annulerade områdena som kantas av lameller .
"Post-cephalothorax" har 5 segment, associerade med 6 somiter med motsvarande par av uniramösa annulerade bihang, som är kantade med lameller. Följande buk är ungefär 60 % av den totala längden, med 6 segment och inga motsvarande ben, som slutar i en gaffelformad svans, i vilken varje flak är sammansatt av tre segment.
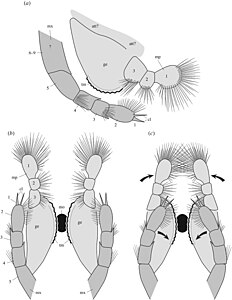
Diagram över underkäken (ljusgrå) och överkäken (mörkgrå) av Waptia i sidled (överst) och underifrån (botten) med de två versionerna som visar bihangens rörelseområde.
Morfologi av cephalothoracic bihang i sidovy (överst) och framifrån

Post-cephalothorax bihang i sidan på vy (a) med närbild av spetsen av bihanget (b) och närbild av lamellerna (c), och en vy underifrån av en sternite med bifogade bihang



Upptäckt


Waptia fieldensis var ett av de första fossilerna som upptäcktes av Charles D. Walcott från Burgess Shale i augusti 1909. En grov skiss av Waptia finns i hans dagbok för den 31 augusti 1909, tillsammans med skisser av Marrella och Naraoia . En formell beskrivning för arten publicerades av Walcott 1912. Arten namngavs efter de två bergen som förbands av den fossila kanten som innehåller Burgess Shale-lokaliteten, Waptaberget och Mount Field of Yoho National Park , British Columbia , Kanada . Namnet på Wapta-berget självt kommer från första nationens Nakoda -ord wapta , som betyder "rinnande vatten"; medan Mount Field fick sitt namn efter den amerikanska telekommunikationspionjären Cyrus West Field .
Tafonomi
Exemplar av Waptia fieldensis återvanns från Burgess Shale Lagerstätte i Kanada , som daterar sig från den mellankambriska perioden (510 till 505 miljoner år sedan). Lokaliteten låg en gång omkring 200 m (660 fot) under vattnet; det var beläget på botten av ett varmt och grunt tropiskt hav intill en undervattens kalkstensklippa (nu Cathedral Limestone Formation ). Undervattensskred orsakade av kollapsen av delar av kalkstensklippan skulle med jämna mellanrum begrava organismerna i området (liksom organismer som bars av jordskreden) i finkornig lera som senare blev skiffer .
Baserat på antalet individer utgör Waptia fieldensis cirka 2,55% av det totala antalet organismer som återvunnits från Burgess Shale och 0,86% av Greater Phyllopod-bädden . Detta gör dem till de tredje vanligaste leddjuren i Burgess Shale (efter Marrella och Canadaspis ). Bara National Museum of Natural History rymmer mer än tusen exemplar av arten från Burgess Shale. Waptia fieldensis hittas ofta disartikulerade, med delar kvar i närheten av varandra.
Exemplar av Waptia , benämnd Waptia jfr. fieldensis har också hittats tillvaratagna från Mellankambriska Spence Shale i Utah , USA. Några av dessa exemplar är förknippade med tredimensionellt bevarade ägg.
Taxonomi
Waptia fieldensis är den enda arten som accepteras under släktet Waptia . Det klassificeras under familjen Waptiidae (etablerat av Walcott 1912).
Vissa författare föreslog att Waptia kan vara allierad med kräftdjur . Andra föreslog att det bara kan vara avlägset besläktat med kräftdjur, åtminstone en medlem av en stamgrupp av kräftdjur, eller till och med av alla leddjur. Waptia omskrevs omfattande 2018 och placerades som en del av kladden Hymenocarina inom Mandibulata , nära besläktad med Crustacea, på grund av den tydliga närvaron av underkäkar och överkäkar.

1975 beskrevs en till synes mycket liknande art från Nedre Kambrium (515 till 520 miljoner år sedan) Maotianshan Shale Lagerstätte i Chengjiang , Kina . Den placerades ursprungligen inom det "ostrakod"-liknande släktet Mononotella , som Mononotella ovata . 1991 omklassificerade Xian-Guang Hou och Jan Bergström det under det nya släktet Chuandianella när ytterligare upptäckter av mer kompletta exemplar gjorde dess likhet med W. fieldensis mer uppenbar. Precis som W. fieldensis hade Chuandianella ovata ett tvåskaligt ryggsköld med en mittkant, ett par stjärtrami, ett enda par antenner och skaftade ögon . 2004 överförde Jun-Yuan Chen preliminärt det till släktet Waptia . C. ovata hade dock åtta abdominala somiter i motsats till fem i W. fieldensis . Dess lemmar var biramösa och var odifferentierade, till skillnad från de av W. fieldensis . Andra författare ansåg att dessa skillnader var tillräckligt för att skilja den från Waptia till sitt eget släkte. 2022 Chuandianella och visade sig sakna mandibler, så den är förmodligen inte nära besläktad med Waptia , trots dess liknande utseende.
återfanns en andra liknande art, Pauloterminus spinodorsalis , från den lägre kambriska Sirius Passet Lagerstätte i Buen-formationen i norra Grönland . Det identifierades också som en möjlig waptiid. Liksom C. ovata hade den biramösa odifferentierade bihang, men den hade också bara fem abdominala somiter som W. fieldensis . Men det dåliga bevarandet av P. spinodorsalis- exemplaren, särskilt av bihangen på huvudet, gör det svårt att fastställa dess taxonomiska placering. Denna svårighet förvärras ytterligare av bevis för att fossilerna av P. spinodorsalis faktiskt kan vara multar ( exuviae ), och inte av det faktiska djuret.
Ekologi

Medan den historiskt sett betraktades som insättningsmatare , som matade genom att sikta genom havsbotten efter ätbara organiska partiklar, ansåg omprövningen 2018 att Waptia hade varit ett aktivt simmande rovdjur av mjukkroppsföremål, med sina första tre par benliknande cephalothorax-bihang. att fånga och manipulera byten, samtidigt som de flyttar sina lamellerade bihang i en rytmisk rörelse för att driva sig själv genom vattnet. Upp och ned rörelse av buken och svansfläkten användes sannolikt för att röra sig vertikalt i vattenpelaren. Den kan ha använt klorna på sina cephalothoracic benliknande bihang för att ibland vila på ytor.
Under 2015 identifierades äggklämmor i sex exemplar från Burgess Shale. Kopplingsstorlekarna var små och innehöll bara upp till 24 ägg, men varje ägg var relativt stort, med en genomsnittlig diameter på 2 mm (0,079 tum). De var fästa längs den inre ytan av den tvåskaliga ryggskölden. Tillsammans med Kunmingella douvillei och Chuandianella från Chengjiang-biotan (cirka 7 miljoner år äldre än Burgess-skiffern) som också hade fossiliserade ägg bevarade inuti ryggskölden, utgör de det äldsta direkta beviset på yngelvård och K-selektion bland djur. Det tyder på att de troligen levde i en miljö som krävde att de vidtog särskilda åtgärder för att säkerställa sina ungas överlevnad.
Se även
externa länkar
- " Waptia fieldensis " . Burgess Shale Fossil Gallery . Kanadas virtuella museum. 2011. Arkiverad från originalet 2020-11-12.
- " Waptia fieldensis ". Smithsonian National Museum of Natural History. 2012.