Överlägsen cervikal ganglion
| Superior cervical ganglion (SCG) | |
|---|---|
 Diagram över den cervikala sympatiskan. (Märkt som "Övre cervikal ganglion")
| |
| Detaljer | |
| Identifierare | |
| latin | ganglion cervicale superius |
| Maska | D017783 |
| TA98 | A14.3.01.009 |
| TA2 | 6608 |
| FMA | 6467 |
| Anatomiska termer för neuroanatomi | |
Det övre cervikala gangliet ( SCG ) är en del av det autonoma nervsystemet (ANS); mer specifikt är det en del av det sympatiska nervsystemet , en uppdelning av ANS som oftast förknippas med fight or flight-svaret . ANS är sammansatt av vägar som leder till och från ganglier , grupper av nervceller. Ett ganglion tillåter en stor mängd divergens i en neuronal bana och möjliggör också en mer lokaliserad krets för kontroll av de innerverade målen. SCG är det enda gangliet i det sympatiska nervsystemet som innerverar huvudet och nacken. Det är den största och mest rostrala (överlägsen) av de tre cervikala ganglierna . SCG innerverar många organ, körtlar och delar av carotissystemet i huvudet.
Strukturera
Plats
SCG är beläget mittemot den andra och tredje halskotan . Den ligger djupt intill höljet på den inre halspulsådern och den inre halsvenen , och framför Longus capitis-muskeln . SCG innehåller neuroner som ger sympatisk innervation till ett antal målorgan i huvudet.
SCG bidrar också till plexus livmoderhalsen . Den cervikala plexusen bildas från en förening av de främre delarna av de fyra övre cervikala nerverna. Var och en får en grå ramus communicans från det överlägsna cervikala gangliet på den sympatiska stammen.
Morfologi och fysiologi och dess neuroner
Den överlägsna cervikala ganglion är rödgrå färg, och vanligtvis formad som en spindel med avsmalnande ändar. Ibland är SCG bred och tillplattad, och ibland sammandragen med intervaller. Den bildades genom sammansmältning av fyra ganglier , motsvarande de fyra övre cervikala nerverna , C1-C4. Kroppen av dessa preganglioniska sympatiska neuroner är specifikt belägna i ryggmärgens laterala horn. Dessa preganglioniska neuroner går sedan in i SCG och synapsar med de postganglioniska neuronerna som lämnar den rostrala änden av SCG och innerverar målorganen i huvudet.
Det finns ett antal neurontyper i SCG, från lågtröskel till högtröskelneuroner. Neuronerna med en låg tröskel har snabbare aktionspotential avfyrningshastighet, medan högtröskelneuronerna har en långsam avfyringshastighet. En annan skillnad mellan SCG-neurontyper görs via immunfärgning . Immunfärgning tillåter klassificering av SCG-neuroner som antingen positiva eller negativa för neuropeptid Y (NPY), som finns i en undergrupp av högtröskelneuroner. NPY-negativa neuroner med låg tröskel är sekretomotoriska neuroner, innerverande spottkörtlar. NPY-negativa neuroner med hög tröskel är vasomotoriska nervceller, som innerverar blodkärl. NPY-positiva neuroner med hög tröskel är vasokonstriktorneuroner, som innerverar iris och tallkottkörteln.
Innervation
SCG tar emot input från ciliospinalcentret . Det ciliospinala centret är beläget mellan C8- och T1-regionerna i ryggmärgen inom den intermediolaterala kolonnen. De preganglioniska fibrerna som innerverar SCG är de bröstryggradsnerverna, som sträcker sig från T1-T8-regionen i ciliospinalcentret. Dessa nerver kommer in i SCG genom den cervikala sympatiska nerven. En mogen preganglionisk axon kan innervera var som helst från 50-200 SCG-celler. Postganglionfibrer lämnar sedan SCG via den interna carotisnerven och den externa carotisnerven. Denna väg för SCG-innervation visas genom stimulering av den cervikala sympatiska nerven, som framkallar aktionspotentialer i både de yttre och interna halsnerverna. Dessa postganglionfibrer skiftar från multipel axoninnervation av sina mål till mindre djupgående multipel axoninnervation eller enkel axoninnervation när SCG-neuronerna mognar under postnatal utveckling.
Fungera
Sympatiskt nervsystem
SCG ger sympatisk innervation till strukturer i huvudet, inklusive tallkottkörteln , blodkärlen i kranialmusklerna och hjärnan, plexus choroid , ögonen, tårkörtlarna, halspulsådern , spottkörtlarna och sköldkörteln . .
Tallkottkörteln
De postganglioniska axonerna i SCG innerverar tallkottkörteln och är involverade i dygnsrytmen . Denna koppling reglerar produktionen av hormonet melatonin, som reglerar sömn- och vakencykler, men påverkan av SCG-neuroninnervation i tallkottkörteln är inte helt klarlagd.
Carotis kropp
De postganglioniska axonerna i SCG innerverar den inre halspulsådern och bildar den inre halspulsådern . Den inre halspulsåden bär de postganglioniska axonerna av SCG till ögat, tårkörteln , slemhinnorna i munnen, näsan och svalget och många blodkärl i huvudet.
Ögat
De postganglioniska axonerna av Superior cervikal ganglion innerverar ögat och tårkörteln och orsakar vasokonstriktion av iris och sclera, pupillutvidgning, utvidgning av palpebralfissuren och minskad produktion av tårar. Dessa svar är viktiga under Fight-or-flight-svaret från ANS. Utvidgning av pupillerna möjliggör en ökad klarhet i synen, och hämning av tårkörteln stoppar tårproduktionen, vilket möjliggör oförsämrad syn och omdirigering av energi någon annanstans.
Hudens blodkärl
De postganglioniska axonerna i SCG innerverar blodkärlen i huden och får kärlen att dra ihop sig. Sammandragning av blodkärlen orsakar ett minskat blodflöde till huden, vilket leder till blekning av huden och kvarhållande av kroppsvärme. Detta spelar in i fight-or-flight-svaret, minskar blodflödet till ansiktshuden och omdirigerar blodet till viktigare områden som musklernas blodkärl.
Vestibulärt system
SCG är kopplat till vestibulära strukturer, inklusive neuroepitel i de halvcirkulära kanalerna och otolitorganen, vilket ger ett tänkbart substrat för modulering av vestibulosympatiska reflexer.
Klinisk signifikans
Horners syndrom
Horners syndrom är en störning som beror på skada på den sympatiska autonoma nervbanan i huvudet. Skador på SCG, en del av detta system, resulterar ofta i Horners syndrom. Skador på T1-T3-regionerna i ryggmärgen är ansvarig för att ögonlocken hänger ( ptos ), förträngning av pupillen ( mios ) och att ögongloben sjunker (synbar Enophthalmos ; inte riktigt sjunken, bara verkar så på grund av den hängande ögonlock). Lesion eller betydande skada på SCG resulterar i en tredje ordningens neuronstörning (se Horners syndrom: Patofysiologi) .
Familjär dysautonomi
Familjär dysautonomi är en genetisk störning som kännetecknas av abnormiteter hos sensoriska och sympatiska neuroner. SCG påverkas avsevärt av denna förlust av neuroner och kan vara ansvarig för några av de resulterande symtomen. I obduktionsstudier är SCG i genomsnitt en tredjedel av normal storlek och har bara 12 procent av det normala antalet neuroner. Defekter i den genetiska kodningen för NGF, som resulterar i mindre funktionell, onormalt strukturerad NGF, kan vara den molekylära orsaken till familjär dysautonomi. NGF är nödvändigt för överlevnad av vissa neuroner så förlust av NGF-funktion kan vara orsaken till neuronal död i SCG.
Historia
Reinnervation
I slutet av 1800-talet upptäckte John Langley att det överlägsna cervikala gangliet är topografiskt organiserat. När vissa områden av det övre cervikala gangliet stimulerades uppstod en reflex i specificerade områden av huvudet. Hans upptäckter visade att preganglioniska neuroner innerverar specifika postganglioniska neuroner. I sina vidare studier av det överlägsna cervikala gangliet upptäckte Langley att det överlägsna cervikala gangliet är regenerativt. Langley skar av SCG ovanför T1-delen, vilket orsakade förlust av reflexer. När de lämnades på egen hand, reinnerverade fibrerna SCG och de initiala autonoma reflexerna återställdes, även om det var begränsad återhämtning av tallkottkörtelfunktionen. När Langley bröt förbindelserna mellan SCG och T1–T5-regionen i ryggmärgen och ersatte SCG med en annan, var SCG fortfarande innerverad i samma del av ryggmärgen som tidigare. När han ersatte SCG med ett T5-ganglion, tenderade gangliet att innerveras av den bakre delen av ryggmärgen (T4–T8). Ersättningen av den ursprungliga SCG med antingen en annan eller en T5-ganglion stödde Langleys teori om topografisk specificitet för SCG.
Forskning
Ganglier i det perifera autonoma nervsystemet används vanligtvis för att studera synaptiska kopplingar. Dessa ganglier studeras eftersom synaptiska kopplingar visar många likheter med det centrala nervsystemet (CNS) och är också relativt tillgängliga. De är lättare att studera än CNS eftersom de har förmågan att växa igen, vilket neuroner i CNS inte har. SCG används ofta i dessa studier och är en av de större ganglierna. Idag studerar neuroforskare ämnen om SCG såsom överlevnad och neuritutväxt av SCG-neuroner, neuroendokrina aspekter av SCG och struktur och vägar för SCG. Dessa studier utförs vanligtvis på råttor, marsvin och kaniner.
Historiska bidrag
- E. Rubin studerade utvecklingen av SCG hos fosterråttor. Forskning om utvecklingen av nerver i SCG har implikationer för den allmänna utvecklingen av nervsystemet.
- Effekterna av ålder på dendritisk arborisering av sympatiska neuroner har studerats i SCG hos råttor. Fynd har visat att det finns betydande dendritisk tillväxt i SCG hos unga råttor men ingen hos åldrade råttor. Hos åldrade råttor fann man att det fanns en minskning av antalet dendriter.
- SCG-celler användes för att studera nervtillväxtfaktor (NGF) och dess förmåga att styra tillväxten av neuroner. Resultaten visade att NGF hade denna styrande, eller tropiska, effekt på neuroner, som styr riktningen för deras tillväxt.
Ytterligare bilder

Den högra sympatiska kedjan och dess förbindelser med bröst-, buk- och bäckenplexus.

Överlägsen cervikal ganglion

Sympatiska anslutningar av ciliar och superior cervikala ganglier.
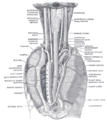
Matstrupens läge och relation i livmoderhalsregionen och i bakre mediastinum. Sett bakifrån.

Den sympatiska bålen och SCG-innerveringen av målorganen i huvudet.





![]() Den här artikeln innehåller text som är allmän egendom från sidan 978 i den 20:e upplagan av Gray's Anatomy (1918)
Den här artikeln innehåller text som är allmän egendom från sidan 978 i den 20:e upplagan av Gray's Anatomy (1918)
- ^ a b Michael J. Zigmond, red. (2000). Fundamental neuroscience (2 uppl.). San Diego: Acad. Tryck. s. 1028–1032 . ISBN 0127808701 .
- ^ Henry Gray. Människokroppens anatomi . 20:e uppl. Philadelphia: Lea & Febiger, 1918 New York: Bartleby.com, 2000. http://www.bartleby.com/107/210.html . Åtkomst 9 juli 2013.
- ^ a b Li, Chen; Horn, John P. (2005). "Fysiologisk klassificering av sympatiska neuroner i råttans överlägsna cervikala ganglion". Journal of Neurophysiology . 95 (1): 187–195. doi : 10.1152/jn.00779.2005 . PMID 16177176 .
- ^ Purves, D; Wigston, DJ (januari 1983). "Neurala enheter i marsvinets övre cervikala ganglion" . Journal of Physiology . 334 (1): 169–78. doi : 10.1113/jphysiol.1983.sp014487 . PMC 1197307 . PMID 6864556 .
- ^ Purnyn, H..; Rikhalsky, O.; Fedulova, S.; Veslovsky, N. (2007). "Sändningsvägar i råttans överlägsna cervikala ganglion". Neurofysiologi . 39 (4–5): 396–399. doi : 10.1007/s11062-007-0053-2 . S2CID 27184650 .
- ^ a b Purves, Dale; Lichtman, Jeff W. (2000). Utveckling av nervsystemet . Sunderland, Mass.: Sinauer Associates. s. 236–238 . ISBN 0878937447 .
- ^ a b Purves, Dale (2012). Neurovetenskap (5 uppl.). Sunderland, Mass.: Sinauer. sid. 465. ISBN 9780878936953 .
- ^ Photoperiodism, melatonin, och tallkottkörteln . London: Pitman Publishing Ltd. 2009. sid. 14.
- ^ Lichtman, Jeff W.; Purves, Dale; Japp, Joseph W. (1979). "I syfte att selektiv innervering av överlägsna cervikala ganglieceller från marsvin" . Journal of Physiology . 292 (1): 69–84. doi : 10.1113/jphysiol.1979.sp012839 . PMC 1280846 . PMID 490406 .
- ^ Pearson, J; Brandeis, L; Goldstein, M (5 oktober 1979). "Tyrosinhydroxylasimmunreaktivitet vid familjär dysautonomi". Vetenskap . 206 (4414): 71–72. Bibcode : 1979Sci...206...71P . doi : 10.1126/science.39339 . PMID 39339 .
- ^ Schwartz, JP; Breakefield, XO (februari 1980). "Förändrad nervtillväxtfaktor i fibroblaster från patienter med familjär dysautonomi" . Proceedings of the National Academy of Sciences of the United States of America . 77 (2): 1154–8. Bibcode : 1980PNAS...77.1154S . doi : 10.1073/pnas.77.2.1154 . PMC 348443 . PMID 6244581 .
- ^ Sanes, Dan H.; Reh, Thomas A.; Harris, William A. (1985). Principer för neural utveckling . San Diego, CA: Academic Press. s. 214–221. ISBN 0-12-300330-X .
- ^ Lingappa, Jaisri R.; Zigmond, Richard E. (2013). "Begränsad återhämtning av pinealfunktion efter regenerering av preganglioniska sympatiska axoner: Bevis för förlust av ganglionisk synaptisk specificitet" . Journal of Neuroscience . 33 (11): 4867–4874. doi : 10.1523/JNEUROSCI.3829-12.2013 . PMC 3640627 . PMID 23486957 .
- ^ Purves, D; Lichtman, JW (oktober 1978). "Bildning och underhåll av synaptiska förbindelser i autonoma ganglier". Fysiologiska recensioner . 58 (4): 821–62. doi : 10.1152/physrev.1978.58.4.821 . PMID 360252 .
- ^ Rubin, E (mars 1985). "Utveckling av råttans överlägsna cervikala ganglion: gangliecellmognad" . Journal of Neuroscience . 5 (3): 673–84. doi : 10.1523/jneurosci.05-03-00673.1985 . PMC 6565020 . PMID 2983044 .
- ^ Andrews, TJ; Li, D; Halliwell, J; Cowen, T (februari 1994). "Effekten av ålder på dendriter i råttans överlägsna cervikala ganglion" . Journal of Anatomy . 184 (1): 111–7. PMC 1259932 . PMID 8157483 .
- ^ Campenot, RB (1977). "Lokal kontroll av neuritutveckling genom nervtillväxtfaktor" . Proc Natl Acad Sci USA . 74 (10): 4516–9. Bibcode : 1977PNAS...74.4516C . doi : 10.1073/pnas.74.10.4516 . PMC 431975 . PMID 270699 .
externa länkar
- Anatomifoto:31:07-0201 vid SUNY Downstate Medical Center
|
Anatomi av det autonoma nervsystemet
| |||||
|---|---|---|---|---|---|
| Huvud |
|
||||
| Nacke |
|
||||
| Bröst |
|
||||
| Buk |
|
||||
| Bäcken |
|
||||
| Myndighetskontroll : Akademiker |
|---|