Cyklinberoende kinaskomplex
Ett cyklinberoende kinaskomplex ( CDKC , cyklin-CDK ) är ett proteinkomplex bildat genom association av en inaktiv katalytisk subenhet av ett proteinkinas, cyklinberoende kinas (CDK), med en regulatorisk subenhet, cyklin . När väl cyklinberoende kinaser binder till cyklin är det bildade komplexet i ett aktiverat tillstånd. Substratspecificiteten för det aktiverade komplexet fastställs huvudsakligen av det associerade cyklinet i komplexet. Aktiviteten hos CDKCs kontrolleras av fosforylering av målproteiner, såväl som bindning av hämmande proteiner.
Struktur och reglering

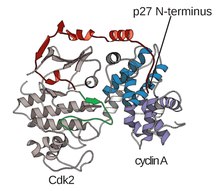
Strukturen av CDKs i komplex med en cyklinsubenhet (CDKC) har länge varit ett mål för struktur- och cellbiologer från och med 1990-talet när strukturen av obundet cyklin A löstes av Brown et al. och samma år Jeffery et al. löste strukturen av humant cyklin A-CDK2-komplex till 2,3 Ångströms upplösning. Sedan denna tid har många CDK-strukturer bestämts till högre upplösning, inklusive strukturerna för CDK2 och CDK2 bundna till en mängd olika substrat, som ses i figur 1. Högupplösta strukturer finns för cirka 25 CDK-cyklinkomplex totalt inom proteinet Databank . Baserat på funktion finns det två allmänna populationer av CDK-cyklinkomplexstrukturer, öppen och sluten form. Skillnaden mellan formerna ligger inom bindningen av cyklinpartners där slutna formkomplex har CDK-cyklinbindning vid både C- och N-ändarna av aktiveringsslingan av CDK, medan de öppna formpartnerna endast binder vid N-änden. Öppna formstrukturer motsvarar oftast de komplex som är involverade i transkriptionsreglering (CDK 8, 9, 12 och 13), medan slutna CDK-cyklinkomplex oftast är involverade i cellcykelprogression och reglering (CDK 1, 2, 6) . Dessa distinkta roller skiljer sig emellertid inte signifikant med sekvenshomologin mellan CDK-komponenterna. I synnerhet, bland dessa kända strukturer tycks det finnas fyra huvudsakliga konserverade regioner: en N-terminal glycinrik loop, en gångjärnsregion, en aC-helix och en T-loopregleringsplats.
Aktiveringsslinga
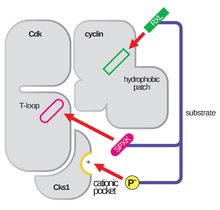
Aktiveringsslingan , även kallad T-loopen, är regionen av CDK (mellan DFG- och APE-motiven i många CDK) som är enzymatiskt aktiv när CDK är bunden till sin funktionsspecifika partner . I CDK-cyklinkomplex består denna aktiveringsregion av en konserverad αL-12 Helix och innehåller en viktig fosforylerbar rest (vanligtvis treonin för CDK-cyklinpartners, men inkluderar även serin och tyrosin) som förmedlar den enzymatiska aktiviteten hos CDK. Det är vid denna essentiella rest (T160 i CDK2-komplex, T177 i CDK6-komplex) som enzymatisk ATP-fosforylering av CDK-cyklinkomplex av CAK (cyklinaktiverande kinas, hänvisar till CDK7-Cyclin H-komplexet i mänskliga celler) äger rum. Efter hydrolysen av ATP för att fosforylera på denna plats kan dessa komplex fullborda sin avsedda funktion, fosforyleringen av cellulära mål. Det är viktigt att notera att i CDK 1, 2 och 6 är T-loopen och en separat C-terminal region de viktigaste platserna för cyklinbindning i CDK, och vilka cykliner som är bundna till var och en av dessa CDK medieras av speciell sekvens av aktiveringsställets T-loop. Dessa cyklinbindningsställen är regionerna med högst variabilitet i CDK trots relativt hög sekvenshomologi som omger αL-12 Helix-motivet för denna strukturella komponent.
Glycinrik region
Den glycinrika loopen (Gly-rik loop) som ses i resterna 12-16 i CDK2 kodar för ett konserverat GXGXXG-motiv över både jäst- och djurmodeller. Den regulatoriska regionen är föremål för differentiell fosforylering vid icke-glycinrester inom detta motiv, vilket gör denna plats föremål för Wee1- och/eller Myt1 -hämmande kinasfosforylering och Cdc25 -defosforylering hos däggdjur. Denna reversibla fosforylering vid den Gly-rika loopen i CDK2 sker vid Y15, där aktiviteten har studerats ytterligare. Studie av denna rest har visat att fosforylering främjar en konformationsförändring som förhindrar ATP- och substratbindning genom sterisk interferens med dessa nödvändiga bindningsställen i aktiveringsslingan av CDK-cyklinkomplexen. Denna aktivitet stöds av den anmärkningsvärda flexibiliteten som den Gly-rika slingan har inom strukturen för de flesta CDK, vilket möjliggör att dess rotation mot aktiveringsslingan har en signifikant effekt på att minska substrataffiniteten utan större förändringar i den övergripande CDK-cyklinkomplexstrukturen.
Gångjärnsregionen
Den konserverade gångjärnsregionen av CDK i eukaryota celler fungerar som en väsentlig brygga mellan den Gly-rika loopen och aktiveringsloopen. CDK kännetecknas av en N-terminal lob som huvudsakligen är vriden beta-ark ansluten via denna gångjärnsregion till en alfahelixdominerad C-terminal lob. I diskussionen om T-loopen och den Gly-rika loopen är det viktigt att notera att dessa regioner, som måste kunna interagera rumsligt för att kunna utföra sina biokemiska funktioner, ligger på motsatta lober av CDK själv. Således förbinder denna gångjärnsregion, som kan variera i längd något mellan CDK-typ och CDK-cyklinkomplex, viktiga regulatoriska regioner av CDK genom att förbinda dessa lober och spelar nyckelroller i den resulterande strukturen av CDK-cyklinkomplex genom att korrekt orientera ATP för enkel katalys av fosforyleringsreaktioner av det sammansatta komplexet.
αC-Helix
αC-Helix-regionen är mycket konserverad över många av däggdjurskinomerna (kinasfamiljen ) . Dess huvudsakliga ansvar är att upprätthålla allosterisk kontroll av det aktiva kinasstället. Denna kontroll manifesterar sig i CDK-cyklinkomplex genom att specifikt förhindra CDK-aktivitet tills dess binder till sin partnerregulator (dvs cyklin eller annat partnerprotein). Denna bindning orsakar en konformationsförändring i αC-Helix-regionen av CDK och gör att den kan flyttas från den aktiva platsklyftan och fullbordar den initiala processen med T-loopaktivering. Med tanke på att denna region är så bevarad över proteinsuperfamiljen av kinaser, har denna mekanism där αC-Helix visat sig vikas ut från kinasets N-terminala lob, vilket möjliggör ökad tillgång till αL-12 Helix som ligger inom T-loopen, anses vara ett potentiellt mål för läkemedelsutveckling.
Cellcykeln
Jästcellscykel
Även om dessa komplex har en mängd olika funktioner, är CDKCs mest kända för sin roll i cellcykeln . Inledningsvis genomfördes studier i Schizosaccharomyces pombe och Saccharomyces cerevisiae (jäst). S. pombe och S. cerevisiae är mest kända för sin association med en enda Cdk, Cdc2 respektive Cdc28, som komplexbinder med flera olika cykliner. Beroende på cyklin påverkas olika delar av cellcykeln. Till exempel, i S. pombe , associerar Cdc2 med Cdk13 för att bilda Cdk13-Cdc2-komplexet. I S. cerevisiae resulterar associationen av Cdc28 med cykliner, Cln1, Cln2 eller Cln3 i övergången från G1-fas till S-fas . Väl i S-fasen dissocierar Cln1 och Cln2 med Cdc28 och komplex mellan Cdc28 och Clb5 eller Clb6 bildas. I G2-fasen resulterar komplex som bildas från associationen mellan Cdc28 och Clb1, Clb2, Clb3 eller Clb4 i progression från G2 - fas till M (mitotisk) fas. Dessa komplex finns också i tidig M-fas. Se tabell 1 för en sammanfattning av jäst CDKCs.
- Tabell 1. CDKCs associerade med cellcykelfaser i jäst
| CDK | Cyclin | Cellcykelfas |
|---|---|---|
| Cdc2 ( S. pombe ) | Cdc13 | G2 till M fasövergång; tidig M-fas |
| Cdc28 ( S. cerevisiae ) | Clnl, Cln2 | G1 till S fasövergång |
| Cdc28 | Clb5, Clb6 | S-fas |
| Cdc28 | Clb1, Clb2, Clb3, Clb4 | G2 till M fasövergång; tidig M-fas |
Från vad som är känt om de komplex som bildas under varje fas av cellcykeln i jäst, har föreslagna modeller framkommit baserade på viktiga fosforyleringsställen och involverade transkriptionsfaktorer.
Däggdjurs cellcykel
Genom att använda informationen som upptäckts genom jästcellcykelstudier har betydande framsteg gjorts när det gäller däggdjurscellcykeln. Det har fastställts att cellcyklerna är likartade och CDKCs, antingen direkt eller indirekt, påverkar cellcykelns fortskridande. Som tidigare nämnts, i jäst, är endast ett cyklinberoende kinas (CDK) associerat med flera olika cykliner. Men i däggdjursceller binder flera olika CDK till olika cykliner för att bilda CDKC. Till exempel associerar Cdk1 (även känd som human Cdc2), den första humana CDK som identifierades, med cykliner A eller B. CyclinA/B-Cdk1-komplex driver övergången mellan G2-fas och M-fas, såväl som tidig M-fas. En annan CDK från däggdjur, Cdk2, kan bilda komplex med cykliner D1, D2, D3, E eller A. Cdk4 och Cdk6 interagerar med cykliner D1, D2 och D3. Studier har visat att det inte finns någon skillnad mellan CDKCs cyklin D1-Cdk4/6, därför kan alla unika egenskaper möjligen kopplas till substratspecificitet eller aktivering. Medan nivåerna av CDK förblir ganska konstanta under hela cellcykeln, fluktuerar cyklinnivåerna. Fluktuationen styr aktiveringen av cyklin-CDK-komplexen och slutligen progressionen genom cykeln. Se tabell 2 för en sammanfattning av CDKC från däggdjursceller som är involverade i cellcykeln.
- Tabell 2. CDKCs associerade med cellcykelfaser i däggdjursceller
| CDK | Cyclin | Cellcykelfas | Icke-cyklinpartnerproteiner |
|---|---|---|---|
| Cdk1 (Cdc2) | Cyclinerna A och B | G2 till M fasövergång; tidig M-fas | Cks1 och Cks2 |
| Cdk2 | cykliner Dl, D2, D3 | G1-fas | KAP, Cks1, p27KIP1 och Spy-1 |
| Cdk2 | Cyclin E | G1 till S fasövergång | KAP, Cks1, p27KIP1 och Spy-1 |
| Cdk2 | Cyclin A | S-fas | KAP, Cks1, p27KIP1 och Spy-1 |
| Cdk4 | cykliner Dl, D2, D3 | G1-fas | HSP90-Cdc37 |
| Cdk6 | cykliner Dl, D2, D3 | G1-fas | p16INK4A, p19INK4D och P18INK4C-cyklin K |
| Cdk8 | Cyclin C | --- | --- |
| Cdk9 | Cyclin T | --- | Tat, AFF4 och TAR |
| Cdk12 | Cyclin K | --- | --- |
| Cdk13 | Cyclin K | --- | --- |

G 1 till S fasprogression
Under sen G 1 -fas binder och fosforylerar CDKC medlemmar av proteinfamiljen retinoblastom (Rb) . Medlemmar av Rb-proteinfamiljen är tumörsuppressorer, som förhindrar okontrollerad cellproliferation som skulle inträffa under tumörbildning. Emellertid tros pRbs också undertrycka de gener som krävs för att övergången från Gi- fas till S-fas ska ske. När cellen är redo att övergå till nästa fas, fosforylerar CDKCs, cyklin D1-Cdk4 och cyklin D1-Cdk6 pRB, följt av ytterligare fosforylering från cyklin E-Cdk2 CDKC. När fosforylering inträffar transkriptionsfaktorer för att irreversibelt inaktivera pRB och progression till S-fasen av cellcykeln följer. Cyklin E-Cdk2 CDKC som bildas i G 1 -fasen hjälper sedan till med initieringen av DNA-replikation under S-fasen.
G 2 till M fasprogression
I slutet av S-fasen är cyklin A associerad med Cdk1 och Cdk2. Under G2-fasen bryts cyklin A ned, medan cyklin B syntetiseras och cyklin B-Cdk1-komplex bildas. Inte bara är cyklin B-Cdk1-komplex viktiga för övergången till M-fasen, utan dessa CDKC spelar en roll i följande regulatoriska och strukturella processer:
- Kromosomal kondensation
- Fragmentering av Golgi-nätverket
- Nedbrytning av nukleär lamina
Inaktivering av cyklin B-Cdk1-komplexet genom nedbrytning av cyklin B är nödvändig för att gå ut ur M-fasen av cellcykeln.
Övrig
Även om majoriteten av de kända CDKCs är involverade i cellcykeln, fungerar inte alla kinaskomplex på detta sätt. Studier har visat att andra CDKC, såsom cyklin k-Cdk9 och cyklin T1-Cdk9, är involverade i replikationsstressresponsen och påverkar transkription . Dessutom kan cyklin H-Cdk7-komplex spela en roll i meios i manliga könsceller och har också visat sig vara involverade i transkriptionsaktiviteter.