Heterocyst

Mikrofotografier av heterocystösa cyanobakterier A–F: Nostoc commune G–H: Nostoc calcicola I–M: Tolypothrix distorta N–R: Scytonema hyalinum Skalstapel = 10 µm , hc, heterocyst, ak, akinete, hm, hormogonium, chrind
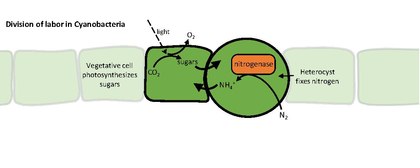
Heterocyster eller heterocyter är specialiserade kvävefixerande celler som bildas under kvävesvältning av vissa trådformiga cyanobakterier , såsom Nostoc punctiforme , Cylindrospermum stagnale och Anabaena sphaerica . De fixerar kväve från dikväve (N 2 ) i luften med hjälp av enzymet nitrogenas , för att förse cellerna i filamentet med kväve för biosyntes.
Nitrogenas inaktiveras av syre, så heterocysten måste skapa en mikroanaerob miljö. Heterocysternas unika struktur och fysiologi kräver en global förändring i genuttryck . Till exempel heterocyster:
- producerar ytterligare tre cellväggar , inklusive en av glykolipid som bildar en hydrofob barriär mot syre
- producerar nitrogenas och andra proteiner involverade i kvävefixering
- bryt ned fotosystem II , som producerar syre
- uppreglera glykolytiska enzymer
- producera proteiner som avlägsnar eventuellt kvarvarande syre
- innehåller polära pluggar som består av cyanofycin som bromsar cell-till-cell-diffusion
Cyanobakterier får vanligtvis ett fixerat kol (kolhydrat) genom fotosyntes . Bristen på vattendelning i fotosystem II hindrar heterocyster från att utföra fotosyntes, så de vegetativa cellerna förser dem med kolhydrater , som tros vara sackaros . De fixerade kol- och kvävekällorna utbyts genom kanaler mellan cellerna i glödtråden. Heterocyster upprätthåller fotosystem I , vilket gör att de kan generera ATP genom cyklisk fotofosforylering .
Enstaka heterocyster utvecklas ungefär var 9-15:e cell, vilket producerar ett endimensionellt mönster längs filamentet. Intervallet mellan heterocyster förblir ungefär konstant även om cellerna i filamentet delar sig. Bakteriefilamentet kan ses som en flercellig organism med två distinkta men ändå beroende celltyper. Sådant beteende är mycket ovanligt hos prokaryoter och kan ha varit det första exemplet på flercellig mönstring i evolutionen . När en heterocyst väl har bildats kan den inte återgå till en vegetativ cell. Vissa heterocystbildande bakterier kan differentiera till sporliknande celler som kallas akineter eller rörliga celler som kallas hormogoni , vilket gör dem till de mest fenotyptiskt mångsidiga av alla prokaryoter.
Genexpression

I miljöer med låg kvävehalt triggas heterocystdifferentiering av den transkriptionella regulatorn NtcA. NtcA påverkar heterocystdifferentiering genom att signalera proteiner som är involverade i processen för heterocystdifferentiering. Till exempel kontrollerar NtcA uttrycket av flera gener inklusive HetR som är avgörande för differentiering av heterocyst. Det är avgörande eftersom det uppreglerar andra gener såsom hetR, patS, hepA genom att binda till deras promotor och därmed fungera som en transkriptionsfaktor . Det är också värt att notera att uttrycket av ntcA och HetR är beroende av varandra och deras närvaro främjar differentiering av heterocyst även i närvaro av kväve. Det har också nyligen upptäckts att andra gener såsom PatA, hetP reglerar differentiering av heterocyst. PatA mönster heterocysterna längs filamenten, och det är också viktigt för celldelning . PatS påverkar heterocystmönstringen genom att hämma heterocystdifferentiering när en grupp differentierande celler samlas för att bilda en pro-heterocyst (omogen heterocyst). Heterocystunderhåll är beroende av ett enzym som kallas hetN. Heterocystbildning hämmas av närvaron av en fixerad kvävekälla, såsom ammonium eller nitrat .
Heterocystbildning
Följande sekvenser äger rum vid bildning av heterocyster från en vegetativ cell:
- Cellen förstoras.
- Granulära inneslutningar minskar.
- Fotosyntetisk lamell omorienterar.
- Väggen blir äntligen treskiktad. Dessa tre lager utvecklas utanför cellens yttre lager.
- Mellanskiktet är homogent.
- Det inre lagret är laminerat.
- Den åldrande heterocysten genomgår vakuolering och bryter slutligen av från filamentet och orsakar fragmentering. Dessa fragment kallas hormogonia (singular hormogonium ) och genomgår asexuell reproduktion.
Cyanobakterierna som bildar heterocyster delas in i ordningarna Nostocales och Stigonematales , som bildar enkla respektive förgrenade filament. Tillsammans bildar de en monofyletisk grupp, med mycket låg genetisk variation .
Symbiotiska relationer
Bakterierna kan också komma in i ett symbiotiskt förhållande med vissa växter. I ett sådant förhållande svarar bakterierna inte på tillgången på kväve, utan på signaler som produceras av växten för differentiering av heterocyst. Upp till 60 % av cellerna kan bli heterocyster, vilket ger fixerat kväve till växten i utbyte mot fixerat kol. Signalen som produceras av växten och det stadium av heterocystdifferentiering den påverkar är okänd. Förmodligen verkar den symbiotiska signalen som genereras av växten före NtcA-aktivering eftersom hetR krävs för symbiotisk heterocystdifferentiering. För den symbiotiska associationen med växten behövs ntcA eftersom bakterierna med muterad ntcA inte kan infektera växter.
Anabaena-Azolla
Ett anmärkningsvärt symbiotiskt förhållande är det mellan Anabaena cyanobakterier och Azolla -växter. Anabaena finns på stjälkarna och inom bladen av Azolla -växter. Azolla - växten genomgår fotosyntes och tillhandahåller fixerat kol för Anabaena att använda som energikälla för dinitrogenaser i heterocystcellerna. I gengäld kan heterocysterna förse de vegetativa cellerna och Azolla -växten med fixerat kväve i form av ammoniak som stödjer tillväxten av båda organismerna.
Detta symbiotiska förhållande utnyttjas av människor i jordbruket. I Asien används Azolla- växter som innehåller Anabaena -arter som biogödsel där kvävet är begränsande såväl som i djurfoder . Olika stammar av Azolla-Anabaena är lämpade för olika miljöer och kan leda till skillnader i växtproduktion. Risgrödor som odlas med Azolla-Anabaena som biogödsel har visat sig resultera i en mycket större kvantitet och kvalitet på produkterna jämfört med grödor utan cyanobakterierna. Azolla-Anabaena- växter odlas före och efter att risgrödor har planterats. När Azolla-Anabaena- växterna växer, ackumulerar de fixerat kväve på grund av verkan av nitrogenasenzymer och organiskt kol från fotosyntesen av Azolla - växterna och Anabaenas vegetativa celler. När Azolla-Anabaena- växterna dör och sönderfaller frigör de stora mängder fixerat kväve, fosfor , organiskt kol och många andra näringsämnen i jorden, vilket ger en rik miljö som är idealisk för odling av risgrödor.
Relationen Anabaena - Azolla har också utforskats som en möjlig metod för att avlägsna föroreningar från miljön, en process som kallas fytoremediation . Anabaena sp. har tillsammans med Azolla caroliniana visat sig vara framgångsrika i att ta bort uran , en giftig förorening orsakad av gruvdrift , samt tungmetallerna kvicksilver (II) , krom(III) och krom(VI) från förorenat avloppsvatten.

Azolla caroliniana växt

Anabaena circinalis filament

Cylindrospermum filament



- ^ Grundläggande biologi (18 mars 2016). "Bakterier" .
- ^ Wolk, CP; Ernst, A.; Elhai, J. (1994). Heterocystmetabolism och utveckling . Cyanobakteriernas molekylära biologi . s. 769–823. doi : 10.1007/978-94-011-0227-8_27 . ISBN 978-0-7923-3273-2 .
- ^ Herrero, Antonia; Muro-Pastor, Alicia M.; Flores, Enrique (15 januari 2001). "Kvävekontroll i cyanobakterier" . Journal of Bakteriologi . 183 (2): 411–425. doi : 10.1128/JB.183.2.411-425.2001 . ISSN 0021-9193 . PMC 94895 . PMID 11133933 .
- ^ Higa, Kelly C.; Callahan, Sean M. (1 augusti 2010). "Ektopiskt uttryck av hetP kan delvis kringgå behovet av hetR i heterocystdifferentiering av Anabaena sp. stam PCC 7120" . Molekylär mikrobiologi . 77 (3): 562–574. doi : 10.1111/j.1365-2958.2010.07257.x . ISSN 1365-2958 . PMID 20545862 .
- ^ Orozco, Christine C.; Risser, Douglas D.; Callahan, Sean M. (2006). "Epistasanalys av fyra gener från Anabaena sp. Stam PCC 7120 föreslår en koppling mellan PatA och PatS i heterocystmönsterbildning" . Journal of Bakteriologi . 188 (5): 1808–1816. doi : 10.1128/JB.188.5.1808-1816.2006 . ISSN 0021-9193 . PMC 1426565 . PMID 16484191 .
- ^ a b lee, Robert Edward. Fykologi (PDF) . Hämtad 9 oktober 2017 .
- ^ Meeks, JC; Elhai, J (2002). "Reglering av celldifferentiering i filamentösa cyanobakterier i frilevande och växtassocierade symbiotiska tillväxtstater" . Mikrobiologi och molekylärbiologi recensioner . 66 (1): 94–121, innehållsförteckning. doi : 10.1128/MMBR.66.1.94-121.2002 . PMC 120779 . PMID 11875129 .
- ^ a b c d van Hove, C.; Lejeune, A. (2002). "Azolla: Anabaena symbios". Biologi och miljö: Proceedings of the Royal Irish Academy . 102B (1): 23–26. doi : 10.1353/bae.2002.0036 . JSTOR 20500136 . S2CID 245843704 .
- ^ a b c d e f Vaishampayan, A.; Sinha, RP; Häder, D.-P.; Dey, T.; Gupta, AK; Bhan, U.; Rao, AL (2001). "Cyanobakteriella biogödselmedel i risjordbruk". Botanisk recension . 67 (4): 453–516. doi : 10.1007/bf02857893 . JSTOR 4354403 . S2CID 20058464 .
- ^ Bocchi, Stefano; Malgioglio, Antonino (2010). "Azolla-Anabaena är ett biogödselmedel för risfält i Po-dalen, ett tempererat risområde i norra Italien" ( PDF) . International Journal of Agronomy . 2010 : 1–5. doi : 10.1155/2010/152158 . ISSN 1687-8159 .
- ^ Singh, S.; Prasad, R.; Singh, BV; Goyal, SK; Sharma, SN (1990-06-01). "Effekt av gröngödsel, blågröna alger och neem-kaka-belagd urea på våtmarksris (Oryza sativa L.)". Jordars biologi och fertilitet . 9 (3): 235–238. doi : 10.1007/bf00336232 . ISSN 0178-2762 . S2CID 38989291 .
- ^ a b Bennicelli, R.; Stępniewska, Z.; Banach, A.; Szajnocha, K.; Ostrowski, J. (2004-04-01). "Azolla carolinianas förmåga att ta bort tungmetaller (Hg(II), Cr(III), Cr(VI)) från kommunalt avloppsvatten". Kemosfär . 55 (1): 141–146. Bibcode : 2004Chmsp..55..141B . doi : 10.1016/j.chemosphere.2003.11.015 . PMID 14720557 .
- ^ Pan, Changchun; Hu, Nan; Ding, Dexin; Hu, Jinsong; Li, Guangyue; Wang, Yongdong (2016-01-01). "En experimentell studie om de synergistiska effekterna mellan Azolla och Anabaena vid avlägsnande av uran från lösningar av Azolla-anabaenas symbiotiska system. " Journal of Radioanalytical and Nuclear Chemistry . 307 (1): 385–394. doi : 10.1007/s10967-015-4161-y . ISSN 0236-5731 . S2CID 82545272 .